
Локализация спинальных и бульбарных вегетативных центров
Центры вегетативной нервной системы разделяют на сегментарные и надсегментарные (высшие вегетативные центры).
Сегментарные центрырасполагаются в нескольких отделах центральной нервной системы, где выделяют 4 очага:
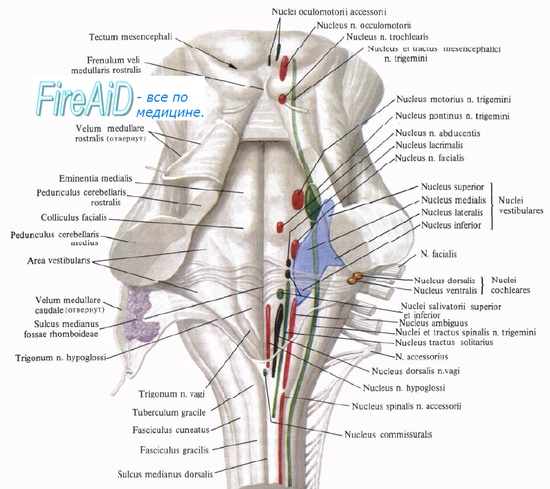
1. Мезенцефалический отдел в среднем мозге - добавочное ядро (Якубовича), nucleus accessorius, и непарное срединное ядро глазодвигательного нерва (III пара).
2. Бульбарный отдел в продолговатом мозге и мосту - верхнее слюноотделительное ядро, nucleus salivatorius superior, промежуточно-лицевого нерва (VII пара), нижнее слюноотделительное ядро, nucleus salivatorius inferior, языкоглоточного нерва (IX пара) и дорсальное ядроблуждающего нерва (X пара), nucleus dorsalis n. vagi.
Оба этих отдела объединяются под названием краниального и относятся к парасимпатическим центрам.
3. Тораколюмбальный отдел - промежуточно-боковые ядра, nuclei intermediolaterales, 16-ти сегментов спинного мозга (С8, Th1-12, L1-3). Они являются симпатическими центрами.
4. Сакральный отдел - промежуточно-боковые ядра, nuclei intermediolaterales, 3-х крестцовых сегментов спинного мозга (S2-4) и относятся к парасимпатическим центрам.
Высшие вегетативные центры (надсегментарные) объединяют и регулируют деятельность симпатического и парасимпатического отделов, к ним относятся:
1. Ретикулярная формация, ядра которой формируют центры жизненно-важных функций (дыхательный и сосудодвигательный центры, центры сердечной деятельности, регуляции обмена веществ и т.д.). Проекция дыхательного центра соответствует средней трети продолговатого мозга, сосудодвигательного центра - нижней части ромбовидной ямки. Нарушение функции ретикулярной формации проявляется вегетативно-сосудистыми расстройствами (кардио-васкулярные, вазомоторные). Кроме того страдают интегративные функции, которые необходимы для формирования целесообразного адаптивного поведения.
2. Мозжечок, принимая участие в регуляции двигательных актов, одновременно обеспечивает эти анимальные функции адаптационно-трофическими влияниями, которые через соответствующие центры приводят к расширению сосудов интенсивно работающих мышц, повышению уровня трофических процессов в последних. Установлено участие мозжечка в регуляции таких вегетативных функций, как зрачковый рефлекс, трофика кожи (скорость заживления ран), сокращение мышц, поднимающих волосы.
3. Гипоталамус - главный подкорковый центр интеграции вегетативных функций, имеет существенное значение в поддержании оптимального уровня обмена веществ (белкового, углеводного, жирового, минерального, водного) и терморегуляции. За счет связей с таламусом он получает разностороннюю информацию о состоянии органов и систем организма, а вместе с гипофизом образует функциональный комплекс - гипоталамо-гипофизарную систему. Гипоталамус в ней выполняет роль своеобразного реле, включающего гипофизарную гормональную цепь в регуляцию различных висцеральных и соматических функций.
4. Особое место занимает лимбическая система обеспечивающая интеграцию вегетативных, соматических и эмоциональных реакций.
5. Полосатое тело имеет ближайшее отношение к безусловнорефлекторной регуляции вегетативных функций. Повреждение или раздражение ядер полосатого тела вызывает изменение кровяного давления, усиление слюно- и слезоотделения, усиление потоотделения.
Высшим центром регуляции вегетативных и соматических функций, а также их координации является кора полушарий большого мозга. Непрерывный поток импульсов от органов чувств, сомы и внутренних органов по афферентным путям поступает в кору головного мозга и через эфферентную часть вегетативной нервной системы, главным образом через гипоталамус, кора оказывает соответствующее влияние на функцию внутренних органов, обеспечивая адаптацию организма к меняющимся условиям окружающей и внутренней среды. Примером кортиковисцеральной связи может служить изменение вегетативных реакций под влиянием словесных сигналов (через вторую сигнальную систему).
Таким образом, вегетативная нервная система, так же как и вся нервная система, построена по принципу иерархии, подчиненности. Схему организации вегетативной иннервации иллюстрирует рис.1.

Рис. 1 Принцип организации вегетативной нервной системы.
Рефлекторная дуга вегетативной
Нервной системы
В простой трехнейронной вегетативной рефлекторной дуге (рис.2), как и в соматической, выделяют те же три звена, а именно: рецепторное, образованное чувствительным (афферентным) нейроном, ассоциативное, представленное вставочным (ассоциативным) нейроном и эффекторное звено, образованное двигательным (эффекторным) нейроном, передающим возбуждение на тот или иной рабочий орган. Нейроны связаны между собой синапсами, в которых с помощью медиаторов происходит передача нервного импульса с одного нейрона на другой.

Рис. 2. Схема рефлекторных дуг соматического (слева) и вегетативного (справа) типов, замыкающихся в спинном мозге.
1- рецептор; 2- чувствительный нейрон спинномозгового ганглия; 3- дорсальный корешок; 4- спинномозговой нерв; 5- вставочный нейрон; 6- двигательный нейрон переднего рога; 7- вентральный корешок; 8- двигательное нервное окончание скелетной мышцы; 9- нейрон симпатического ядра бокового рога; 10- преганглионарное волокно; 11- белая соединительная ветвь; 12- периферический вегетативный ганглий; 13- эффекторный нейрон; 14- постганглионарное волокно; 15- серая соединительная ветвь; 16- двигательное нервное окончание на гладкой мышце; 17и 18- волокна пирамидного пути.
Чувствительные нейроны представлены псевдоуниполярными клетками спинномозгового узла, так же как и в соматической нервной системе. Их периферические отростки заканчиваются рецепторами в органах. Поэтому информация о состоянии органов растительной и животной жизни стекается в спинномозговые узлы, и в этом смысле они являются смешанными соматически-вегетативными узлами. Центральный отросток чувствительного нейрона в составе заднего корешка вступает в спинной мозг и нервный импульс переключается на вставочный нейрон, клеточное тело которого расположено в боковых рогах (латерально-промежуточное ядро тораколюмбального или сакрального отделов) серого вещества спинного мозга.
Вставочный нейрон отдает аксон, который покидает спинной мозг в составе передних корешков и достигает одного из вегетативных узлов, где вступает в контакт с эффекторным (двигательным) нейроном.
Таким образом, второе звено вегетативной рефлекторной дуги отличается от соматической, во-первых, местом локализации тела вставочного нейрона, во-вторых, протяженностью и положением аксона, который в отличие от соматической нервной системы никогда не остается в пределах спинного мозга. Еще большие различия в строении третьего звена рефлекторной дуги. В отличие от соматической рефлекторной дуги, где двигательные нейроны расположены в передних рогах спинного мозга, для вегетативной рефлекторной дуги характерно расположение двигательного нейрона за пределами центральной нервной системы - в вегетативных узлах, аксоны которых направляются к рабочему органу, а это значит, что весь эфферентный путь подразделяется на два участка: предузловой (преганглионарный) - аксон вставочного нейрона и послеузловой (постганглионарный) - аксон двигательного нейрона вегетативного узла. Таким образом, в вегетативной рефлекторной дуге эфферентный периферический путь двухнейронный.
В простой трехнейронной вегетативной рефлекторной дуге, замыкающейся в пределах мозгового ствола, тело первого нейрона располагается в чувствительных узлах черепных нервов, второго - в вегетативных ядрах черепных нервов (мезенцефалический и бульбарный отделы) и третьего - в вегетативных узлах.
Достигая эффекторов (гладких мышц и желез), нервные импульсы вызывают сокращение мышцы или изменение секреторной деятельности железы, что в свою очередь вызывает раздражение рецепторов этих органов и отсюда поток импульсов по афферентным волокнам направляется обратно в ядра спинного или головного мозга, неся ежемоментную информацию о состоянии данного органа. Наличие обратной связи (обратной афферентации), с одной стороны позволяет осуществлять контроль за правильностью исполнения команд, с другой - вносить дополнительную своевременную коррекцию в выполнении ответной реакции организма.
Таким образом, в основе строения и функции вегетативной нервной системы, как и соматической, лежит замкнутая кольцевая цепь рефлексов, которая способствует наиболее полному приспособлению организма к окружающей среде.
ВЕГЕТАТИВНЫЕ УЗЛЫ
Узлы вегетативной нервной системы устроены однотипно, но отличаются друг от друга своей локализацией. По этому топографическому признаку их делят условно на три группы (порядка).Узлы I порядка, околопозвоночные (паравертебральные), ganglia trunci sympathici, образуют симпатический ствол, расположенный по сторонам позвоночного столба. Узлы II порядка, предпозвоночные (превертебральные) или промежуточные, ganglia intermedia, расположены впереди позвоночника, входят в состав вегетативных сплетений и также как и узлы I порядка относятся к симпатическому отделу вегетативной нервной системы. Узлы III порядка составляют конечные (терминальные) узлы, ganglia terminalia. Они в свою очередь разделяются на околоорганные и внутриорганные и относятся к парасимпатическим узлам.
Различная степень удаленности вегетативных узлов от спинного мозга или мозгового ствола, с одной стороны, и рабочего органа, с другой, сказывается на длине преганглионарных и постганглионарных волокон. Наибольшую длину имеют преганглионарные волокна, связанные с узлами III порядка. Постганглионарные волокна этих узлов, наоборот, очень короткие. Обратная картина наблюдается при рассмотрении волокон, связанных с узлами I порядка. Здесь преганглионарные волокна короткие, а постганглионарные - длинные. Преганглионарные волокна покрыты миелиновой оболочкой, благодаря чему имеют белый цвет. Их диаметр чаще составляет 2-3,5 мкм, а скорость распространения импульсов колеблется от 3 до 18 м/сек. Постганглионарные волокна лишены миелина, серого цвета, диаметр - до 2 мкм, скорость проведения импульса - 1-3 м/сек.
Снаружи вегетативный узел покрыт соединительнотканной капсулой, от которой внутрь отходят тяжи, между которыми располагаются группы клеток. Тело каждого нейрона имеет тонкую соединительнотканную капсулу. Между ней и телом нейрона - клетки сателлиты, которые выполняют опорную, защитную и трофическую функции.
В узлах выделяют три типа нейронов:
1. Клетки Догеля первого типа - мультиполярные с множеством коротких ветвящихся дендритов и одним длинным аксоном, направляющимся к рабочему органу (рис. 3). Это двигательные нейроны.
2. Клетки Догеля второго типа - чувствительные, мультиполярные с длинными дендритами, которые покидают узел и заканчиваются рецепторами на гладкомышечных или железистых клетках. Аксон может заканчиваться здесь же в ганглии, или направляться в соседние.
3. Клетки Догеля третьего типа представляют местные ассоциативные элементы, соединяющие своими отростками несколько клеток I и II типов как своего узла, так и соседних узлов.
 | Рис.3. Различные типы нейронов соматической (а) и вегетативной (б) частей нервной системы (из А.Г.Кнорре и И.Д.Лев). 1- пирамидная клетка коры головного мозга; 2- клетка Пуркинье из коры мозжечка; 3- клетка-зерно из коры мозжечка; 4- двигательная клетка переднего рога спинного мозга; 5- биполярная клетка сетчатки; 6- униполярная клетка сетчатки; 7- клетка 1 типа Догеля; 8- клетка II типа Догеля; 9- клетка промежуточно-латерального ядра бокового рога спинного мозга. |
Вегетативные узлы выполняют следующие функции:
1. Передачу нервного импульса с преганглионарной на постганглионарную часть эфферентного пути. Эта функция обусловлена наличием в узле двигательных нейронов.
2. Рефлекторную или замыкательную. Благодаря наличию не только двигательных , но и чувствительных нейронов в вегетативных узлах могут замыкаться периферические рефлекторные дуги, что превращает их в периферические нервные центры, способные осуществлять местные рефлексы на основе принципа саморегуляции.
3. Рецепторную, так как в узлах имеются рецепторы, которыми заканчиваются дендриты чувствительных нейронов спинномозговых и черепных узлов. Благодаря этим центростремительным связям состояние и деятельность вегетативных узлов находятся под контролем центральной нервной системы.
4. Через вегетативный узел проходят транзитно афферентные и эфферентные волокна.
Кроме анатомически обособленных ганглиев, по ходу вегетативных ветвей периферических нервов встречается большое количество нервных клеток, мигрировавших сюда в ходе эмбрионального развития.
Последнее изменение этой страницы: 2016-08-15; Нарушение авторского права страницы
Центры регуляции вегетативных функций
Центры регуляции вегетативных функций разделяются на спинальные, стволовые (бульбарные, мезэнцефалические), гипоталамические, мозжечковые, центры ретикулярной формации, лимбической системы, корковые. В основе их взаимодействия лежит принцип иерархии. Каждый более высокий уровень регуляции обеспечивает и более высокую степень интеграции вегетативных функций.
На уровне спинного мозга происходит регуляция просвета зрачка, величины глазной щели, сосудистого тонуса, потоотделения. Стимуляция этих центров приводит к усилению и учащению сердечной деятельности, расширению бронхов. Здесь расположены также центры дефекации, мочеиспускания, половых рефлексов (эрекции и эякуляции).
Эти центры находятся в продолговатом мозге, мосту, среднем мозге.
За счет ядер блуждающих нервов происходит торможение деятельности сердца, возбуждение слезоотделения, усиление секреции слюнных, желудочных желез, поджелудочной железы, желчевыделения, усиление сокращений желудка и тонкой кишки.
Сосудодвигательный центр отвечает за рефлекторное сужение и расширение сосудов и регуляцию кровяного давления.
Дыхательный центр регулирует смену вдоха и выдоха.
В продолговатом мозге находятся центры, с помощью которых осуществляются такие сложные рефлексы, как сосание, жевание, глотание, чихание, кашель, рвота.
В передних буграх четверохолмия в среднем мозге располагаются центры, регулирующие зрачковый рефлекс и аккомодацию глаза.
Гипоталамус является главным подкорковым центром интеграции висцеральных процессов, что обеспечивается вегетативными, соматическими и эндокринными механизмами.
Стимуляция ядер задней группы гипоталамуса сопровождается реакциями, аналогичными раздражению симпатической нервной системы: расширение зрачков и глазных щелей, учащение сердечных сокращений, сужение сосудов и повышение АД, торможение моторной активности желудка и кишечника, увеличение содержания в крови адреналина и норадреналина, концентрации глюкозы. Задняя область гипоталамуса отвечает за регуляцию теплопродукции и оказывает тормозящее влияние на половое развитие.
Стимуляция передних ядер гипоталамуса приводит к эффектам, подобным раздражению парасимпатической нервной системы: сужение зрачков и глазных щелей, замедление частоты сердечных сокращений, снижение артериального давления, усиление моторной активности желудка и кишечника, увеличение секреции желудочных желез, стимуляция секреции инсулина и снижение уровня глюкозы в крови. Передние ядра регулируют теплоотдачу и оказывают стимулирующее влияние на половое развитие.
Средняя группа ядер гипоталамуса обеспечивает регуляцию метаболизма и водного баланса. Вентромедиальные ядра отвечают за насыщение, латеральные ядра – за голод (центры голода и насыщения). Паравентрикулярное ядро – центр жажды.
Гипоталамус отвечает за эмоциональное поведение, формирование половых и агрессивно-оборонительных реакций.
С помощью нейротропных средств можно избирательно воздействовать на гипоталамические структуры и регулировать состояние голода, жажды, аппетита, страха, половые реакции.
Центры лимбической системы.
Эти центры отвечают за формирование вегетативного компонента эмоциональных реакций, пищевое, сексуальное, оборонительное поведение, регуляцию систем, обеспечивающих сон и бодрствование, внимание.
Благодаря наличию активирующего и тормозного механизмов мозжечок может оказывать стабилизирующее влияние на деятельность висцеральных органов посредством корригирования висцеральных рефлексов.
Центры ретикулярной формации.
Ретикулярная формация осуществляет тонизирование и повышение активности других вегетативных нервных центров.
Центры коры больших полушарий.
Кора больших полушарий осуществляет высший интегративный контроль вегетативных функций посредством нисходящих тормозных и активирующих влияний на ретикулярную формацию и другие подкорковые вегетативные центры. Координирует вегетативные и соматические функции в системе поведенческого акта.
Бульбарный уровень регуляции кровообращения. Бульбарный сосудодвигательный центр. Вазомоторный центр.
Современные представления о бульбарных механизмах регуляции кровообращения основаны на исследованиях, в которых показано влияние как перерезок на разных уровнях, так и стимуляции различных бульбарных структур на артериальное давление и частоту сердечных сокращений, а также на импульсации в симпатических и парасимпатических нервах. Раздражение продолговатого мозга и варолиева моста оказывает либо прес-сорные, либо депрессорные изменения АД в зависимости от локализации раздражающих электродов: прессорные зоны располагаются, как правило, ростральнее и латеральнее депрессорных. При перерезке на уровне ядер лицевого нерва отделяется расположенная ростральнее часть прессорнои области, тогда как депрессорная остается интактной. В этом случае снижается артериальное давление и уменьшается импульсация в эфферентных вегетативных нервах. Перерезка продолговатого мозга на уровне задвижки ведет к еще большему снижению давления и полному прекращению импульсации по вегетативным нервам. Бульбарная медиальная депрессорная область оказывает угнетающее тоническое влияние на спинальные пре-ганглионарные нейроны, способные к спонтанной импульсивной активности.
Указанный медуллярный комплекс, реализующий влияния на сердечнососудистую систему через симпатические нервы, называют бульбарным сосудодвигательным центром. Функционально он включает в себя и структуры дорсального ядра блуждающего нерва, обеспечивающие тонические кардиоингибирующие влияния. Поэтому говорят о едином бульбарном центре сердечно-сосудистой системы, получающем информацию от различных рецепторных зон и обеспечивающем поддержание циркуляторного гомеостазиса. Это достигается благодаря реципрокному взаимодействию симпатических и парасимпатических рефлекторных влияний на сердце, а также дозировкой тонических констрикторных влияний симпатических нервов на сосуды.
Доказаны проекции нейронов каудальной части вентральных отделов продолговатого мозга к клеткам его ростральной части, что свидетельствует о возможности тонического угнетения активности этих клеток. Функционально значимы связи структур вентральных отделов продолговатого мозга с ядром солитарного тракта, которое играет ключевую роль в обработке афферентации от хемо- и барорецепторов сосудов.
К вентральным отделам продолговатого мозга информация поступает как от его структур — ядер солитарного тракта, area postrema, вестибулярных ядер, ядер шва, обоюдного ядра, так и от клеток спинного мозга и супрабульбарных структур центральной нервной системы. Наиболее значимыми в функциональном плане для структур ростральной части вентральных отделов являются их связи с симпатическими преганглионарными нейронами спинного мозга, а для структур каудальных вентролатеральных отделов — восходящие проекции к нейросекреторным клеткам гипоталамуса.
Структуры ростральной части вентральных отделов продолговатого мозга играют ключевую роль в тоническом контроле уровня артериального давления. Стимуляция нейронов рострального вентролатерального ядра вызывает подъем АД, тахикардию, торможение барорецепторных рефлексов, высвобождение адреналина из надпочечников и аргинин-вазопрессина из нейросекреторных клеток гипоталамуса. Возбуждение нейронов каудальной части вентральных отделов ведет к снижению системного АД. Структуры вентральных отделов участвуют в контроле сердечной деятельности и через обоюдное ядро блуждающего нерва.
Нейроны ростральной части вентральных отделов продолговатого мозга являются звеном дуг собственных и сопряженных сосудистых рефлексов, а нейроны каудальной части — модулируют реализацию на сосуды указанных рефлексов.
Сердечно-сосудистый центр продолговатого мозга оказывает тоническое возбуждающее влияние на преганглионарные симпатические нейроны тораколюмбального отдела спинного мозга. Однако его влияние является не единственными связями, конвергирующими на спинальных вегетативных нейронах. В медуллярной области происходит переключение путей нисходящих возбуждающих и тормозных кардиоваскулярных влияний, исходящих из вышележащих отделов центральной нервной системы. Собственная тоническая активность спинальных нейронов в норме зависит от нисходящих бульбарных и супрабульбарных влияний, эти нисходящие влияния полностью доминируют над их спонтанной активностью. Несмотря на то что сегментарные спинальные вазомоторные рефлексы (например, побледнение кожи над областью внутрибрюшинной воспалительной реакции) могут иметь место, замыкание собственных сердечно-сосудистых рефлексов происходит не ниже, чем на бульбарном уровне. Однако и бульбарный уровень регуляции в естественных условиях модулируется сложной совокупностью нисходящих к нему влияний, среди которых ведущую роль играют нейрогуморальные влияния гипоталамического происхождения.
Центры ВНС расположены в спинном и головном мозге. Их следует представлять как согласованно действующие ансамбли нейронов, ответственных за выполнение той или иной функции. Вегетативные центры разделяют на высшие (надсегментарные) и низшие (сегментарные). Координирующее влияние сегментарных центров распространяется на отдельные функции и осуществляется через определенные нервы. Надсегментарные центры контролируют деятельность сегментарных вегетативных центров, осуществляют их интеграцию с центрами соматической нервной системы и другими регулирующими системами – эндокриннной, кровеносной и т. д.
Сегментарные вегетативные нервные центры образованы телами нейронов, которые по своему положению в рефлекторной дуге являются вставочными.
По функции выделяют симпатические и парасимпатические вегетативные центры.
По топографии различают центры головного мозга (краниальные) и спинного мога (спинальные).
В отличие от строго сегментарного расположения соматических (анимальных) центров, для вегетативных нервных центров характерна очаговость. Имеются четыре таких очага:
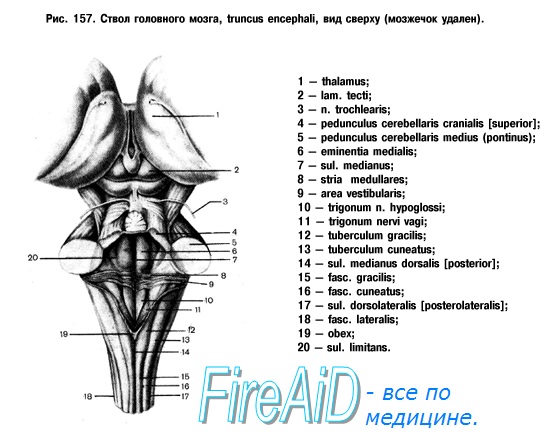
1. мезенцефалический (парасимпатический) – добавочное ядро III пары черепных нервов, nucl. accessorius.
2. Понто-бульбарный (парасимпатический) – верхнее и нижнее слюноотделительные ядра VII и IX пары – nucl. salivatorius superior, nucl. salivatorius inferior, и вегетативное ядро X пары – nucl. dorsalis n. vagi.
Оба эти очага являются краниальными.
3. Тораколюмбальный (симпатический) – в боковых рогах спинного мозга (nucl. intermediolaterales) на протяжении сегментов С8, Th1-L2.
4. Сакральный (парасимпатический) – nucl. parasympathici sacrales, в сером веществе сегментов S2-S4.
Перечисленные очаги, или сегментарные вегетативные центры, находятся под контролирующим и коррегирующим воздействием надсегментарных (высших) центров, которые расположены в стволе мозга, мозжечке, подкорковых структурах и в коре полушарий головного мозга. Эти центры не являются специализированными (симпатическими или парасимпатическими), а объединяют в себе регуляцию обоих отделов вегетативной нервной системы. Так, в стволе головного мозга существенную роль в регуляции вегетативных функций играет ретикулярная формация, formatio reticularis, (около 100 ядер), ядра которой формируют дыхательный, сосудодвигательный, пищеварительный центры. В мозжечке – центры, регулирующие трофику кожи, сосудодвигательные рефлексы, сокращение мышц, поднимающих волосы, mm. arrectores pili. Важную роль в обеспечении вегетативных функций отводят гипоталамической области. Здесь сосредоточены центры, ответственные за поддержание постоянства внутренней среды организма (гомеостаз). Благодаря наличию обширных нервных и сосудистых связей между гипоталамусом и гипофизом, обе эти структуры объединяют в единую гипоталамо-гипофизарную систему, осуществляющую нейро-гуморальную регуляцию деятельности всех органов растительной жизни, желез внутренней секреции. В подкорковых базальных ядрах (corpus striatum et corpus amygdaloideum) содержатся центры терморегуляции, слюно- и слезоотделения.
Особое место среди высших вегетативных центров занимает лимбическая система. Это – структуры среднего, промежуточного и конечного мозга (сводчатая извилина, миндалевидное тело, мозговая полоска таламуса, гипоталамус, гиппокамп, свод, прозрачная перегородка и др.). все эти структуры объединяют в общее понятие – висцеральный мозг, в который поступает весь поток сенсорной информации и на базе ее первичного синтеза формируются определенные биологические потребности – мотивации, обеспечивается эмоциональная окраска как вегетативных так и соматических реакций организма.
И, наконец, корковые вегетативные центры, которые сосредоточены преимущественно в лобных и теменных долях и осуществляют объединение (интеграцию) вегетативных и анимальных функций всего организма.
Как отмечалось выше, в основе взаимоотношения вегетативных центров лежит принцип иерархии – объем регулирующего влияния тем больше, чем выше положение центра, причем действие высших центров реализуется не только через низшие, но и через другие регулирующие системы – эндокринную, кровеносную (например, гипоталамо-гипофизарная система).
Центры вегетативной нервной системы разделяют на сегментарные и надсегментарные (высшие вегетативные центры).
Сегментарные центры располагаются в нескольких отделах центральной нервной системы, где выделяют 4 очага:
Мезенцефалический отдел в среднем мозге - добавочное ядро (Якубовича) глазодвигательного нерва (III пара).
Бульбарный отдел в продолговатом мозге и мосту: 1) верхнее слюноотделительное ядропромежуточно-лицевого нерва (VII пара), 2) нижнее слюноотделительное ядро языкоглоточного нерва (IX пара) и 3) дорсальное ядро блуждающего нерва (X пара.
Оба этих отдела относятся к парасимпатическим центрам.
Тораколюмбальный отдел - промежуточно-боковые ядра 16-ти сегментов спинного мозга от 8-го шейного до 3-го поясничного включительно (Ш8, Г1-12, П1-3). Они являются симпатическими центрами.
Сакральный отдел - промежуточно-боковые ядра 3-х крестцовых сегментов спинного мозга от 2-го до 4-го включительно (К2-4) и относятся к парасимпатическим центрам.
Высшие вегетативные центры (надсегментарные) объединяют и регулируют деятельность симпатического и парасимпатического отделов, к ним относятся:
1. Ретикулярная формация, ядра которой формируют центры жизненно-важных функций (дыхательный и сосудодвигательный центры, центры сердечной деятельности, регуляции обмена веществ и т.д.).
2. Мозжечок, в котором имеются трофические центры.
Гипоталамус - главный подкорковый центр интеграции вегетативных функций, имеет существенное значение в поддержании оптимального уровня обмена веществ (белкового, углеводного, жирового, минерального, водного) и терморегуляции.
Полосатое тело имеет ближайшее отношение к безусловнорефлекторной регуляции вегетативных функций. Повреждение или раздражение ядер полосатого тела вызывает изменение кровяного давления, усиление слюно- и слезоотделения, усиление потоотделения.
Высшим центром регуляции вегетативных и соматических функций, а также их координации является кора полушарий большого мозга.
Вегетативная Рефлекторная дуга
Вегетативная нервная система , как и соматическая нервная система, реализует свои функции по принципу рефлексов.
В простой вегетативной рефлекторной дуге, как и в соматической, выделяют три звена, а именно: 1) рецепторное, образованное чувствительным (афферентным) нейроном, 2) ассоциативное, представленное вставочным нейроном и 3) эффекторное звено, образованное двигательным (эфферентным) нейроном, передающим возбуждение на рабочий орган.
Нейроны связаны между собой синапсами, в которых с помощью медиаторов происходит передача нервного импульса с одного нейрона на другой.
Чувствительные нейроны (I нейрон) представлены псевдоуниполярными клетками спинномозгового узла. Их периферические отростки заканчиваются рецепторами в органах. Центральный отросток чувствительного нейрона в составе заднего корешка вступает в спинной мозг и нервный импульс переключается на вставочный нейрон, клеточное тело которого расположено в боковых рогах (латерально-промежуточное ядро тораколюмбального или сакрального отделов) серого вещества спинного мозга (II нейрон).
Аксон вставочного нейрона покидает спинной мозг в составе передних корешков и достигает одного из вегетативных узлов, где вступает в контакт с двигательным нейроном (III нейрон).
Таким образом, вегетативная рефлекторная дуга отличается от соматической, во-первых, местом локализации вставочного нейрона ( в боковых рогах, а не в задних), во-вторых, протяженностью и положением аксона вставочного нейрона, который в отличие от соматической нервной системы выходит за пределы спинного мозга, в-третьих, тем, что двигательный нейрон расположен не в передних рогах спинного мозга, а в вегетативных узлах (ганглиях), а это значит, что весь эфферентный путь подразделяется на два участка: предузловой (преганглионарный) - аксон вставочного нейрона и послеузловой (постганглионарный) - аксон двигательного нейрона вегетативного узла.
Узлы вегетативной нервной системы по топографическому признаку делят условно на три группы (порядка).
Узлы I порядка, околопозвоночные, образуют симпатический ствол, расположенный по сторонам позвоночного столба.
Узлы II порядка, предпозвоночные или промежуточные, расположены впереди позвоночника, входят в состав вегетативных сплетений. Узлы I и II порядка относятся к симпатическому отделу вегетативной нервной системы.
Узлы III порядка составляют конечные узлы. Они в свою очередь разделяются на околоорганные и внутриорганные и относятся к парасимпатическим узлам.
Преганглионарные волокна покрыты миелиновой оболочкой, благодаря чему имеют белый цвет. Постганглионарные волокна лишены миелина, серого цвета.
В узлах выделяют три типа нейронов:
Клетки Догеля первого типа - двигательные нейроны.
Клетки Догеля второго типа - чувствительные нейроны. Благодаря наличию чувствительных клеток в узле, рефлекторные дуги могут замыкаться через вегетативный узел - периферические рефлекторные дуги.
Клетки Догеля третьего типа представляют ассоциативные нейроны.
РАЗЛИЧИЯ ВЕГЕТАТИВНОЙ И СОМАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Вегетативная нервная система отличается от соматической следующими признаками:
Вегетативная нервная система иннервирует гладкую мускулатуру и железы, и кроме того она обеспечивает трофическую иннервацию всех тканей и органов, включая скелетную мускулатуру, т.е.иннервирует все органы и ткани, а соматическая иннервирует только скелетную мускулатуру.
Важнейший отличительный признак вегетативного отдела - это очаговый характер расположения центров (ядер) в стволе головного мозга (мезенцефалический и бульбарный отделы) и спинном мозге (тораколюмбальный и сакральный отделы). Соматические же центры располагаются в пределах центральной нервной системы равномерно (сегментарно).
Различия в строении рефлекторной дуги (см. выше).
Деятельность вегетативной нервной системы основана не только на центральных рефлекторных дугах, но и на периферических, двухнейронных, замыкающихся в вегетативных узлах.
Вегетативная нервная система обладает избирательной чувствительностью к гормонам. Это обусловлено тем, что переключение импульса в синапсах осуществляется с помощью химического вещества - медиатора.
Читайте также:
