
Мышечные веретена чувствительны к растяжению мышц
Функции мышечных веретен
Как подсказывает название " рецептор растяжения ", мышечное веретено реагирует на растяжение мышцы. Рис. 38.3 демонстрирует изменение активности афферентного аксона при переходе мышечного веретена от укороченного состояния (без нагрузки; во время сокращения экстрафузальных волокон ) к состоянию удлинения при растяжении мышцы. Сокращение заставляет мышечное веретено укорачиваться, поскольку оно лежит параллельно экстрафузальным волокнам.
Активность афферентов мышечных веретен зависит от механического растяжения афферентных окончаний на интрафузальных волокнах . При сокращении экстрафузальных волокон мышечное волокно укорачивается, расстояние между витками афферентного нервного окончания уменьшается и частота разряда в афферентном аксоне падает. И наоборот, когда вся мышца подвергается растяжению, мышечное веретено тоже удлиняется (потому что его концы прикреплены к соединительнотканной сети внутри мышцы) и растягивание афферентного окончания повышает частоту его импульсного разряда.
Частота разряда в афферентах групп Iа и II пропорциональна длине мышечного веретена; это заметно как во время линейного растяжения ( рис. 38.4 , слева), так и при расслаблении мышцы после растяжения ( рис. 38.4 , справа). Такая реакция называется статическим ответом афферентов мышечного веретена. Однако первичные и вторичные афферентные окончания отвечают на растяжение по-разному.
Первичные окончания чувствительны и к степени растяжения, и к его скорости, тогда как вторичные реагируют преимущественно на величину растяжения ( рис. 38.4 ). Эти различия определяют характер активности окончаний двух типов. Частота разряда первичного окончания достигает максимума во время растяжения мышцы, а при расслаблении он прекращается. Реакция такого типа называется динамическим ответом афферентных аксонов группы Iа ( рис. 38.4 , средний ряд записей). Постукивание по мышце (либо по ее сухожилию) или синусоидальное растяжение более эффективно вызывают разряд в первичном афферентном окончании, чем во вторичном.
Судя по характеру ответов, первичные афферентные окончания сигнализируют как о мышечной длине, так и о скорости ее изменения, а вторичные афферентные окончания передают информацию только о длине. Эти различия зависят в основном от разницы в механических свойствах интрафузальных волокон с ядерными сумкой и цепочкой. Первичные и вторичные окончания есть на волокнах обоих типов, тогда как вторичные находятся преимущественно на волокнах с ядерной цепочкой. Средняя (экваториальная) часть волокна с ядерной сумкой лишена сократительных белков из-за скопления клеточных ядер, поэтому легко растягивается. Однако сразу после растяжения она стремится вернуться к своей исходной длине, хотя концевые части волокна удлиняются. Феномен, который называется "оползание", обусловлен вязко-упругими свойствами этого интрафузального волокна. В результате наблюдается вспышка активности первичного окончания с последующим ослаблением активности до нового статического уровня частоты импульсов.
В отличие от волокон с ядерной сумкой у волокон с ядерной цепочкой длина изменяется в более близком соответствии с изменениями длины экстрафузальных мышечных волокон, потому что средняя их часть содержит сократительные белки. Следовательно, вязко-упругие характеристики волокна с ядерной цепочкой более однородны, оно не подвержено оползанию и его вторичные афферентные окончания генерируют только статические ответы.
До сих пор мы рассматривали поведение мышечных веретен только в отсутствие активности гамма-мотонейронов . Вместе с тем их эфферентная иннервация чрезвычайно значима, поскольку определяет чувствительность мышечных веретен к растяжению. Например, на рис. 38.5 ,а представлена активность афферента мышечного веретена во время постоянного растяжения. Как уже говорилось, при сокращении экстрафузальных волокон ( рис. 38.5 ,б) веретена перестают испытывать нагрузку и разряд их афферентов прекращается. Однако влиянию разгрузки противодействует эффект стимуляции гамма-мотонейронов. Такая стимуляция заставляет мышечное веретено укорачиваться вместе с экстрафузальными волокнами ( рис. 38.5 ,в). Точнее, укорачиваются только два конца мышечного веретена; срединная (экваториальная) его часть, где находятся клеточные ядра, не сокращается из-за отсутствия сократительных белков. В результате срединная часть веретена удлиняется, так что афферентные окончания растягиваются и возбуждаются. Этот механизм очень важен для нормальной деятельности мышечных веретен, так как в результате нисходящих двигательных команд от головного мозга происходит, как правило, одновременная активация альфа-мотонейронов и гамма-мотонейронов и, следовательно, сопряженное сокращение экстрафузальных и интрафузальных мышечных волокон ( рис. 38.6 ).
Другой способ влияния афферентов на рефлекторную активность - через их взаимодействие с интрафузальными волокнами с ядерной сумкой и с ядерной цепочкой.
Существует два типа гамма-мотонейронов : динамические и статические ( рис. 38.2 ). Динамические двигательные гамма-аксоны оканчиваются на интрафузальпых волокнах с ядерной сумкой, а статические - на волокнах с ядерной цепочкой. При активации динамического гамма-мотонейрона усиливается динамический ответ афферентов группы Iа ( рис. 38.7 ,г), а при активации статического гамма-мотонейрона возрастают статические ответы афферентов обеих групп - Iа и II ( рис. 38.7 ,в), и одновременно может снижаться динамический ответ. Разные нисходящие пути оказывают предпочтительное влияние на динамические либо статические гамма- мотонейроны, изменяя таким образом характер рефлекторной активности спинного мозга .
Мышечные веретена состоят из интрафузальных мышечных волокон, количество которых не превышает 12. (В данном контексте экстрафузалъные мышечные волокна считают обычными). Крупные интрафузальные волокна начинаются от полюсов (концов) мышечных веретен и фиксируются к соединительной ткани — перимизию. Некрупные интрафузальные волокна фиксируются к коллагеновой капсуле веретена. В средней части (экваторе) веретена саркомеры практически полностью замещены ядрами, расположенными в форме сумки—сумчато-ядерные мышечные волокна (характерно для широких волокон) или цепочки — цепочечно-ядерные мышечные волокна (характерно для тонких волокон).
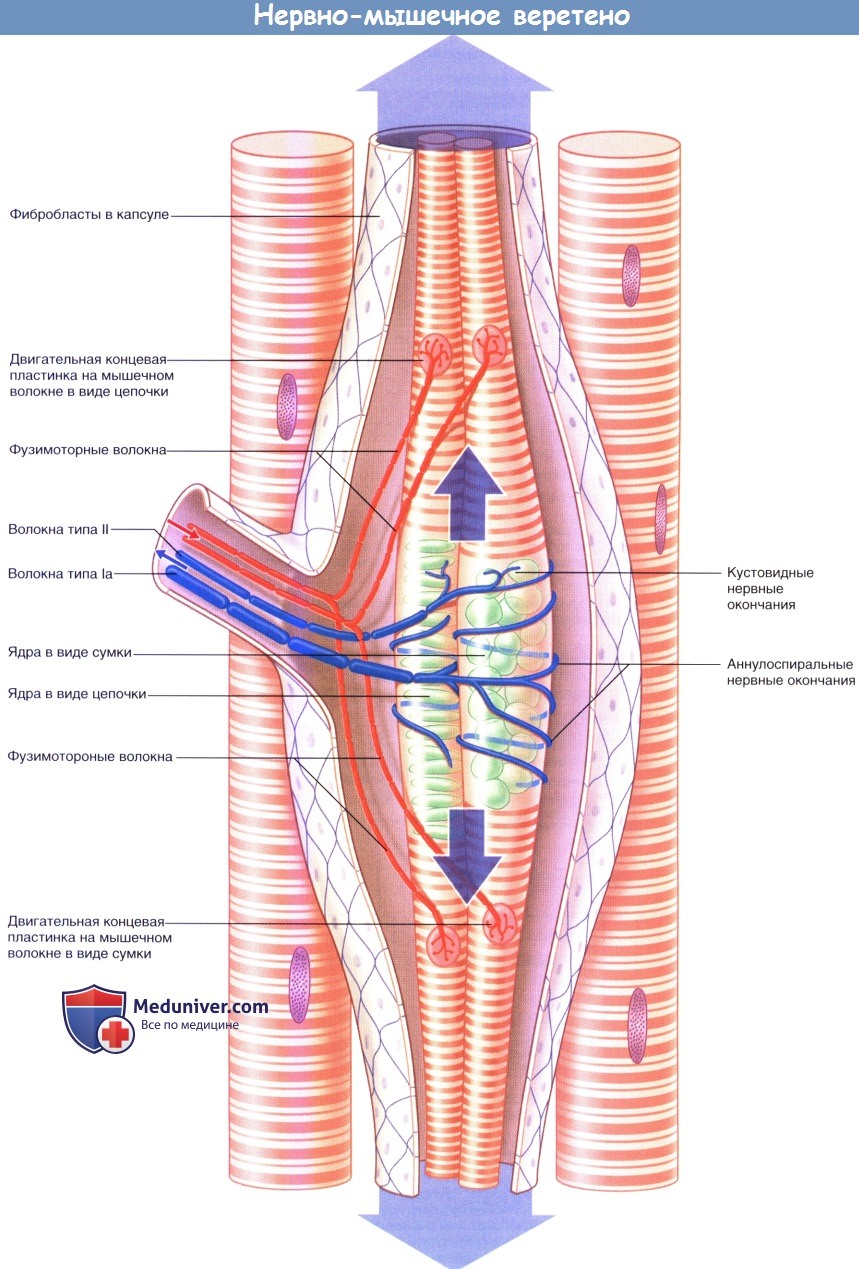
Упрощенное строение нервно-мышечного веретена. Большими стрелками обозначено пассивное растяжение аннулоспиральных нервных окончаний при удлинении мышцы в состоянии покоя.
Стрелками среднего размера обозначено активное растяжение аннулоспиральных нервных окончаний фузимоторными нервными волокнами.
Активное растяжение в достаточной степени компенсирует эффект разгрузки одновременного сокращения экстрафузальных мышечных волокон.
Стрелками маленького размера показаны направления проведения импульсов к мышечному веретену и от него при сокращении мышцы.
1. Иннервация. Иннервацию мышечных веретен осуществляют двигательные и чувствительные нервные волокна. Двигательные нервные волокна — фузимоторные—в соответствии с толщиной относят к типу Аγ, а нервные волокна экстрафузальных мышц — к типу Аα. Фузимоторные аксоны разветвляются и иннервируют поперечно-исчерченные сегменты концевых участков интрафузальных мышц. Единичные первичные чувствительные нервные волокна типа 1а оборачиваются спирально-кольцевым образом вокруг сумчато-ядерных или цепочечно-ядерных мышечных волокон. Вторичные кустовидные чувствительные нервные окончания образованы нервными волокнами II типа и располагаются с одной или с двух сторон от первичного волокна.
2. Активация. Мышечные веретена представляют собой рецепторы растяжения. При растяжении мышечного веретена ионные каналы поверхностной мембраны чувствительных нервных окончаний открываются, за счет чего создаются положительные электроволны. В результате их суммирования в области терминального участка аксона чувствительного нервного волокна происходит образование рецепторного потенциала, который при достижении пороговых значений генерирует нервные импульсы.
Растяжение мышечных веретен может происходить пассивным и активным путями.
3. Пассивное растяжение. Пассивное растяжение мышечного веретена происходит при пассивном удлинении всего мышечного брюшка. Например, при воспроизведении сухожильных рефлексов, в частности коленного, во время удара по сухожилию происходит пассивное растяжение мышечных веретен брюшка четырехглавой мышцы бедра. Волокна типов Iа и II направляются к спинному мозгу и образуют синапсы с дендритами α-мотонейронов. (Принадлежность мотонейронов к типу а определяется Аα-диаметром их аксонов.)
Сухожильные рефлексы относят к моносинаптическим. Для них характерен латентный период (промежуток времени до ответной реакции на стимул), составляющий 15-25 мс.
Помимо возбуждающего действия на гомонимные мотонейроны (т. е. двигательные нейроны, иннервирующие одни и те же мышцы), афферентные волокна мышечных веретен опосредованно через вставочные тормозные мотонейроны оказывают тормозное действие на а-мотонейроны мышц-антагонистов. Этот процесс получил название реципрокного торможения. Задействованные тормозные нейроны называют вставочными нейронами типа Iа.
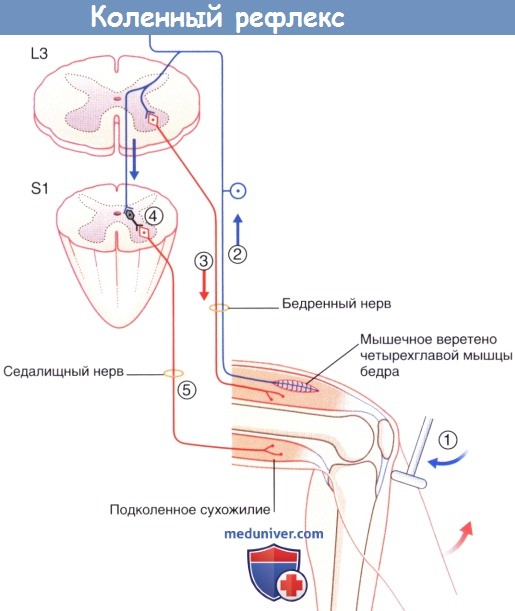
Коленный рефлекс, в котором задействовано реципрокное торможение.
Стрелочками указано проведение нервных импульсов.
(1) Удар по коленной связке вызывает растяжение мышечных веретен в четырёхглавой мышце спины.
(2) Мышечные веретена посылают возбуждающий потенциал в спинной мозг.
(3) α-Мотонейроны вызывают подергивание четырехглавой мышцы и разгибание колена.
(4 и 5) Тормозные вставочные нейроны типа Ia полностью подавляют активность подколенного сухожилия.
4. Кодирование информации. Первичные афференты мышечных веретен проявляют наибольшую активность во время растяжения. Они определяют скорость растяжения мышцы: чем быстрее происходит растяжение, тем больше импульсов они генерируют.
Вторичные афференты мышечных веретен по сравнению с первичными активнее задействованы в процессе поддержания определенной позиции и определяют степень растяжения мышцы: чем выше степень статического растяжения, тем больше формируется импульсов.
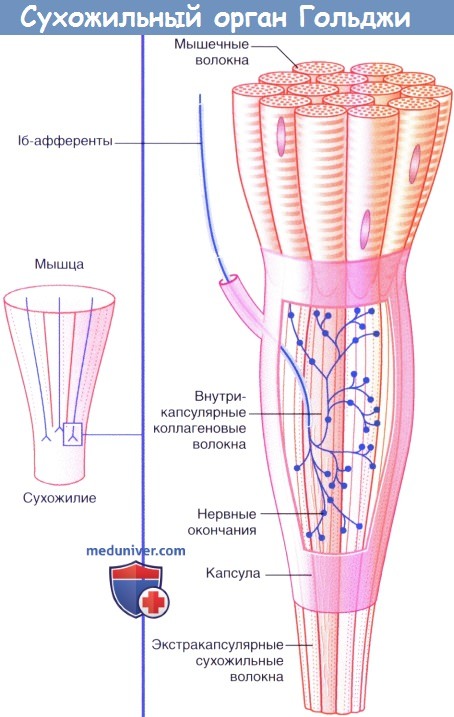
б) Нервные окончания сухожилий. Сухожильные органы Гольджи располагаются в местах соединения сухожилий с мышцами. Единичное нервное волокно типа Ib распадается на сложноорганизованные мельчайшие разветвления, оплетающие пучки сухожильных волокон, связанные с соединительно-тканной капсулой.
Десятки мышечных волокон погружаются в сухожильные интракапсулярные волокна, которые последовательно связываются с другими мышечными волокнами в пределах определенной мышцы. Под действием натяжения, возникающего в процессе сокращения мышцы, происходит активация луковицеобразных нервных окончаний. В связи с тем, что скорость возникновения импульсов в исходном нервном волокне зависит от силы натяжения, сухожильные нервные окончания определяют силу сокращения мышцы.
Сухожильный орган Гольджи. 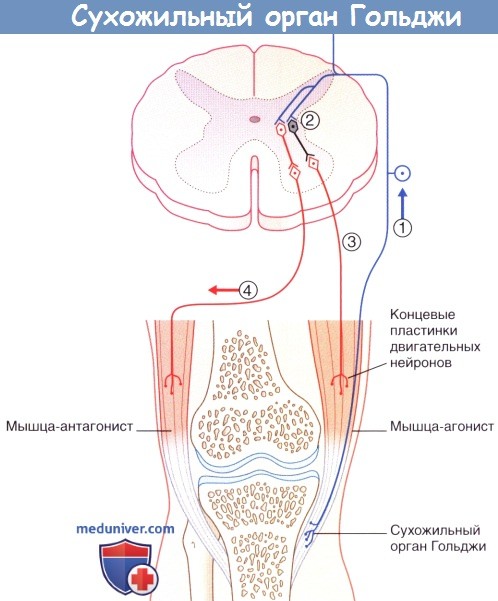
Рефлекторные эффекты при стимуляции сухожильного органа Гольджи.
(1) Сокращение мышцы-агониста вызывает возбуждение афферентов сухожильного органа Гольджи,
что приводит к (2) возбуждению тормозных вставочных нейронов, образующих синапсы с гомонимными мотонейронами,
а также (3) возбуждению тормозных вставочных нейронов, образующих синапсы с (4) мотонейронами мышцы-антагониста.
в) Свободные нервные окончания. В мышцах имеется множество свободных нервных окончаний, большинство которых расположено во внутримышечной соединительной ткани и покрывающих мышцы фасциях. Эти нервные окончания отвечают за болевую чувствительность, возникающую при непосредственном повреждающем воздействии или при накоплении продуктов распада, к которым относят молочную кислоту.
г) Иннервация суставов. Немиелинизированные нервные волокна со свободными нервными окончаниями в большом количестве присутствуют в связках и капсулах суставов, а также во внешних частях внутрисуставных менисков. Эти нервные волокна обеспечивают болевую чувствительность при деформациях суставов, а также участвуют в формировании защитного рефлекса для капсулы сустава. Так, например, переднюю капсулу запястья иннервируют срединный и локтевой нервы; внезапное растяжение при насильственном разгибании вызывает рефлекторную активацию двигательных волокон, что приводит к сгибанию верхней конечности в лучезапястном суставе.
В экспериментах на животных показано, что при воспалении сустава в возбужденном состоянии находится большее количество нервных волокон, чем при растяжении капсулы здорового сустава. Вероятно, существуют нервные окончания, единственным стимулом которых служит воспалительный процесс.
Инкапсулированные нервные окончания, расположенные внутри суставных капсул и вокруг них, представлены тельцами Руффини, реагирующими на натяжение, ламеллярными окончаниями, воспринимающими давление, а также тельцами Пачини, ответственными за ощущение вибрации.
д) Миофасциальный болевой синдром. Миофасциальный болевой синдром—распространенное заболевание, проявляющееся региональной мышечной болезненностью, связанной с чрезмерной чувствительностью пучков напряженных мышечных волокон. (Сходные синдромы наблюдают при другом заболевании—фибромиалгии. Однако при фибромиалгии происходит центральное нарушение болевой чувствительности—дисфункция системы модуляции боли.)
Прикосновение к пучкам мышечных волокон с повышенной чувствительностью вызывает боль; клинически эту область называют миофасциальной триггерной точкой. Для боли не характерно распространение в области дерматома какого-то определенного нерва; в некоторых случаях боль может выходить за пределы триггерной точки—иррадиировать. Кроме того, боль могут сопровождать вегетативные проявления, такие как покраснение и пилоэрекция. Возникновение триггерных точек может быть связано с травмой мышц, чрезмерными нагрузками во время профессиональной деятельности или спортивных занятий при нарушении процесса нормального восстановления.
Спонтанно активирующиеся очаги получили название активных миофасциальных триггерных точек (МТТ), а находящиеся в данный момент времени в неактивном состоянии—латентных миофасциальных триггерных точек. Происходящие процессы недостаточно ясны с точки зрения патофизиологии, однако установлено, что тканевая жидкость, окружающая активные МТТ, содержит большее количество ассоциированных с воспалением веществ (например, брадикинины, простагландины, протоны Н + ).
Со временем боль может захватывать новые области или усиливаться в результате раздражения нейронов задних рогов. Высвобождение другими ветвями раздраженных нейронов субстанции Р может привести к возникновению новых МТТ в области той же или прилежащей мышцы.
Стойкое сокращение мышечных волокон, прилежащих к узелковым утолщениям, объясняют инактивацией ацетилхолинэстеразы в базальной мембране их концевых двигательных пластинок. Принципы лечения этого заболевания—длительное пассивное растяжение пораженных мышц, длительное давление в горизонтальном положении больного (например, путем подкладывания теннисного мячика под пораженную область), а также механическое повреждение прокалыванием иглой или введением местных анестетиков и/или стероидов.
е) Резюме. Мышцы. Двигательная единица состоит из двигательного нейрона и иннервируемой им группы мышечных волокон. Каждая двигательная единица содержит определенный гистохимический вид мышечных волокон. Концевое утолщение нервно-мышечного синапса, содержащее пузырьки с АХ, отделено от синаптических складок сарколеммы базальной мембраной, в которой присутствует ацетилхолинэстераза.
В состав мышечных веретен входят интрафузальные мышечные волокна, активация которых происходит с обоих концов за счет γ-фузимоторных нейронов. Чувствительные волокна типа Iа образуют первичные аннулоспиральные нервные окончания в области средней части (экватора), а волокна типа II — вторичные нервные окончания. Оба типа волокон являются рецепторами растяжения. Растяжение может происходить пассивным (например, при сухожильном рефлексе) или активным путем во время фузимоторной активности. Гомонимные мотонейроны—моносинаптические; мышцы-антагонисты реципрокно ингибируются посредством вставочных нейронов Iа. Первичные афференты мышечных веретен определяют скорость мышечного сокращения, а вторичные — степень.
В процессе произвольных движений происходит одновременная активация А α- и Аγ-мотонейронов.
Сухожильные органы Гольджи определяют силу мышечных сокращений. В состав сухожильных органов входит инкапсулированное сухожилие, иннервацию которого осуществляют афференты типа Iб, вызывающие двухсинаптическое торможение гомонимных мотонейронов и реципрокное возбуждение мышц-анта-гонистов.
Свободные внутримышечные нервные окончания обеспечивают болевую чувствительность.
Суставы. Свободные нервные окончания в большом количестве присутствуют в связках и капсулах суставов, а также во внешних частях внутрисуставных менисков. Они обеспечивают болевую чувствительность и формирование суставного защитного рефлекса. Инкапсулированные нервные окончания принимают импульсы от движений суставов.
- Вернуться в оглавление раздела "Неврология."
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
Сегодня мы поговорим о функциональной анатомии человека и подробно рассмотрим нервно-мышечное веретено. Но перед этим, давайте вначале поймем, что это.
Если мы обратимся к Wikipedia, там указано, что в каждой поперечнополосатой мышце содержатся мышечные веретена. Мышечные веретена, в соответствии с названием, имеют форму веретена длиной несколько миллиметров и диаметром несколько десятых долей миллиметра. Веретена расположены в толще мышцы параллельно обычным мышечным волокнам.
Проприоцепция - это общее понимание положения тела относительно друг друга. Она служит в качестве важного механизма обеспечения безопасности движения тела. Кроме того, это адаптивная система организации для мышц и суставов. Нервная система взаимодействует с мышцами, сухожилиями и суставами с помощью различных проприорецепторов, что позволяет чувствовать любое изменение в положении тела.

Нервно-мышечное веретено - это проприорецептор, который распространяется по всей скелетной мышечной ткани и отслеживает изменения в длине ткани. Мышцы веретена включают в себя специализированные скелетные мышечные волокна называемые интрафузальными волокнами. Они окружены катушкой чувствительных нервных окончаний. Чувствительные нервы, или афферентные волокна контролируют скорость и величину растяжения в мышцах.
Если появляется сильное растяжение или достаточно сильное, чтобы потенциально повредить ткани, альфа-эфферентные волокна воздействуют на окружающие экстра фузальные волокна, чтобы сократить мышцы, тем самым защищая их от вредного перерастяжения. Этот ответ называется миотатическим рефлексом. Если вы когда-либо проверяли у врача проверить свои рефлексы, вы стали свидетелями миотатического рефлекса. Удар молоточком по связке надколенника на передней части колена обычно побуждает четырехглавую мышцу на передней части бедра к быстрому сокращению. Ваша нога как бы выпрыгивает, что говорит о том, что ваши нервно-мышечные веретена работают правильно. Все скелетные мышцы содержат мышечные веретена и защищены миотатическим рефлексом.
Очень важно понимать взаимосвязь между скоростью движения и активацией мышечных веретен. Повторяющиеся растяжение активирует мышечные веретена, что вызывает
сокращение мышцы. Кроме того, повторная активация мышечных веретен будет создавать повышенную чувствительность к растяжению, активизируя рефлекс растяжения мышцы при меньшем воздействии. Это создает повышенное напряжение мышц и агрессивную реакцию мышечных веретен, которые ограничивают подвижность и создают дискомфорт и дисфункцию мышцы.
Десенсибилизация или уменьшение уровня чувствительности мышечных веретен является одной из стратегий избавления от гипертонуса мышц и улучшение активного диапазона движений. Мышечные веретена становятся менее чувствительными, когда они полностью контролируют движения. Обеспечение пассивных движений или участие в активных движениях, которые медленно и мягко удлиняют мышечную ткань, будет настраивать мышечные веретена и уменьшать ограничения нервно-мышечной подвижности. Растяжение должно быть медленным и достаточно мягким, чтобы убедить мышечные веретена, что вы движетесь достаточно аккуратно, чтобы не вызвать травму.
При применении таких техник, как шведский массаж или других методов, которые включают в себя изменение положения тела клиента, не забывайте про мышечные веретена. Быстрое или чрезмерное удлинение мышц может спровоцировать спазм мышц и гипертонус. Если цель состоит в том, чтобы улучшить подвижность и удлинить ткани, сначала убедитесь, что вы безопасно и аккуратно работаете с мышечными веретенами. Делайте движения медленно и дайте время проприорецепторам понять, что происходит и осознать безопасность движения. После того, как мышцы привыкнут, вы можете растягивать мышечные ткани без помех.
В каждой поперечнополосатой мышце содержатся мышечные веретена. Мышечные веретена, в соответствии с названием, имеют форму веретена длиной несколько миллиметров и диаметром несколько десятых долей миллиметра. Веретена расположены в толще мышцы параллельно обычным мышечным волокнам. Мышечное веретено имеет соединительнотканную капсулу. Капсула обеспечивает механическую защиту элементов веретена, расположенных в полости капсулы, регулирует химическую жидкую среду этих элементов и этим обеспечивает их взаимодействие. В полости капсулы мышечного веретена расположено несколько особых мышечных волокон, способных к сокращению, но отличающихся от обычных мышечных волокон мышцы как по строению, так и по функции. Эти мышечные волокна, расположенные внутри капсулы, назвали интрафузальными мышечными волокнами (лат.: intra — внутри; fusus — веретено); обычные мышечные волокна называются экстрафузальными мышечными волокнами (лат.: extra — вне, снаружи; fusus — веретено). Интрафузальные мышечные волокна тоньше и короче экстрафузальных мышечных волокон. Выделяют два главных типа интрафузалъных мышечных волокон.
Один тип интрафузального мышечного волокона — волокно с ядерной сумкой (bursa nuclearis)Это волокно имеет в средней части около сотни компактно собранных клеточных ядер. Утолщенная средняя часть такого интрафузального волокна представляет собой сумку с ядрами. Именно потому эти волокна назвали сумчато-ядерными.
Другой тип волокна, волокно с ядерной цепочкой (vinculun nucleare) , имеет ядра распределенные в виде цепочки по длинной оси интрафузального волокна. Именно потому эти волокна называют цепочно-ядерными интрафузальными мышечными волокнами. Цепочечно-ядерные волокна вдвое тоньше и почти вдвое короче, чем сумчато-ядерные волокна.
Иннервация мышечных веретен
В полость каждого мышечного веретена на уровне ядерной зоны проникают нервные волокна и кровеносные сосуды. Среди нервных волокон одно — толстое миелинизированное нервное волокно. Внутри мышечного веретена одиночное крупное волокно ветвится и посылает терминали дендритов к каждому интрафузальному мышечному волокну любых разновидностей. Конечные нервные ветви обвиваются вокруг средних частей интрафузальных волокон, образуя так называемое аннулоспиральное окончание (лат.: anulus — колечко, завиток; anulo- в форме кольца). Афферентные нервные волокна, образующие аннулоспиральные окончания, принадлежат к типу Iа. Их называют первичными афферентными волокнами мышечных веретен. В соответствии с этим аннулоспиральные нервные окончания носят название первичных чувствительных (сенсорных) окончаний. Полагают, что каждое мышечное веретено иннервируется только одним афферентным нервным волокном типа Iа.
Экстрафузальные и интрафузальные мышечные волокна иннервируются разными типами моторных нейронов (эфферентных нервных волокон). Экстрафузальные волокна иннервируются α-мотонейронами, а интрафузальные — γ-мотонейронами. Тела γ-мотонейронов, посылающих по своим аксонам (фузимоторные нервные волокна) управляющие сигналы к мышечным веретенам, значительно меньше по размерам, чем тела α-мотонейронов, управляющих экстрафузальными мышечными волокнами. Фузимоторные нервные волокна значительно тоньше эфферентных волокон, управляющих сокращениями мышц. Фузимоторные нервные волокна называют γ-(эфферентными) нервными волокнами. В пределах мышцы γ-волокна разветвляются и иннервируют несколько мышечных веретен. Внутри каждого веретена γ-волокна иннервируют несколько интрафузальных мышечных волокон. γ-волокна образуют несколько типов окончаний на периферических (полярных) участках интрафузальных мышечных волокон. Эти окончания называют γ-концевыми пластинками, если они локализованы на сумчато-ядерных интрафузальных мышечных волокнах, а также γ-кустовидными нервными окончаниями, если они локализованы на цепочечноядерных интрафузальных мышечных волокнах. γ-концевые пластинки подобны обычным концевыми пластинкам, расположенным на экстрафузальных мышечных волокнах). γ-кустовидные окончания представляют собой длинные тонкие структуры, разветвленные в виде диффузной сети. Каждое γ-волокно образует только один тип терминалей: либо только кустовидные окончания, либо только концевые пластинки.
Приходящие по эфферентным нервным окончаниям импульсы вызывают сокращение мышечных волокон, что ведёт к деформации афферентных окончаний и приводит к возникновению нервных импульсов, частота которых пропорциональна скорости изменения длины мышечных волокон и величине этого изменения.
• Мышечные веретена — рецепторы растяжения мышц, бывают двух типов:
o с ядерной сумкой
o с ядерной цепочкой
• Сухожильный орган Гольджи — рецепторы сокращения мышц. При сокращении мышцы сухожилие растягивается и его волокна пережимают рецепторное окончание, активируя его.
Нервно-мышечное веретено — сложный рецептор, который включает видоизмененные мышечные клетки, афферентные и эфферентные нервные отростки и контролирует как скорость, так и степень сокращения и растяжение скелетных мышц.
Строение мышечных веретен
В каждой поперечнополосатой мышце содержатся мышечные веретена. Мышечные веретена, в соответствии с названием, имеют форму веретена длиной несколько миллиметров и диаметром несколько десятых долей миллиметра. Веретена расположены в толще мышцы параллельно обычным мышечным волокнам. Мышечное веретено имеет соединительнотканную капсулу. Капсула обеспечивает механическую защиту элементов веретена, расположенных в полости капсулы, регулирует химическую жидкую среду этих элементов и этим обеспечивает их взаимодействие. В полости капсулы мышечного веретена расположено несколько особых мышечных волокон, способных к сокращению, но отличающихся от обычных мышечных волокон мышцы как по строению, так и по функции. Эти мышечные волокна, расположенные внутри капсулы, назвали интрафузальными мышечными волокнами (лат.: intra — внутри; fusus — веретено); обычные мышечные волокна называются экстрафузальными мышечными волокнами (лат.: extra — вне, снаружи; fusus — веретено). Интрафузальные мышечные волокна тоньше и короче экстрафузальных мышечных волокон. Выделяют два главных типа интрафузалъных мышечных волокон.
Один тип интрафузального мышечного волокона — волокно с ядерной сумкой (bursa nuclearis)Это волокно имеет в средней части около сотни компактно собранных клеточных ядер. Утолщенная средняя часть такого интрафузального волокна представляет собой сумку с ядрами. Именно потому эти волокна назвали сумчато-ядерными.
Другой тип волокна, волокно с ядерной цепочкой (vinculun nucleare) , имеет ядра распределенные в виде цепочки по длинной оси интрафузального волокна. Именно потому эти волокна называют цепочно-ядерными интрафузальными мышечными волокнами. Цепочечно-ядерные волокна вдвое тоньше и почти вдвое короче, чем сумчато-ядерные волокна.
Сухожильный орган Гольджи (нервно-сухожильное веретено) — рецепторный орган, располагающийся в местах соединения мышечных волокон с коллагеновыми пучками сухожилий.
Весь орган окружён тонкой соединительнотканной капсулой. Нервное волокно, подходящее к веретену утрачивает миелиновый слой и, распадаясь на терминальные ветви, проходит между пучками коллагеновых волокон сухожилия.
Сокращаясь, мышца растягивает сухожилие. Это вызывает возбуждение нернвых окончаний чувствительных нейронов спинного мозга. Они возбуждают тормозные нейроны спинного мозга, которые тормозят соответствующие двигательные нейроны, что предотвращает перерастяжения мышцы. Известно, что порог возбуждения рецептеров сухожильного органа Гольджи при механическом растяжении мышцы выше, чем аналогичный порог возбуждения мышечных веретен. Это позволяет мышце сокращаться без всяких помех до определенного предела
20.
Моно- и полисинаптические рефлексы спинного мозга. Миотатический и обратный миотатический рефлекс. 21. моно и полисинаптические рефлексы спинного мозга. Миотатический и обратный миотатический рефлекс.
Спинной мозг — часть центральной нервной системы, расположенная в позвоночном канале.
Рефлекс (от лат reflexus – отраженный) - стереотипная реакция организма на определенное воздействие, проходящая с участием нервной системы. Рефлексы существуют у многоклеточных живых организмов, обладающих нервной системой. По степени сложности нейронной организации рефлекторные дуг различают моносинаптические, дуги которых состоят из афферентного и эфферентного нейронов (например, коленный), и полисинаптические, дуги которых содержат также 1 или несколько промежуточных нейронов и имеют 2 или несколько синаптических переключений (например, флексорный).
Существуют и более сложные рефлекторные дуги, включающие один или неск. вставочных нейронов между афферентной и эфферентной частями рефлекса. Примером простейшего полисинаптического (более чем с одним синапсом) рефлекса может служить сухожильный рефлекс. Сенсорный концевой орган — тельца Гольджи — находится в сухожилиях. Увеличение нагрузки на сухожилие, вызываемое обычно сокращением прикрепленной к нему мышцы, и есть возбуждающий раздражитель, к-рый приводит к растяжению телец Гольджи и возникновению в них импульсной активности, распростр. по соотв. афферентному волокну. Идущий от сухожильного сенсорного концевого органа афферент заканчивается на вставочном нейроне в спинном мозге. Этот вставочный нейрон оказывает тормозящее действие на α-мотонейрон, понижая активность в его эфферентном аксоне. Поскольку этот аксон возвращается к мышце, прикрепленной к растянутому сухожилию, мышца расслабляется и нагрузка на сухожилие снижается. Рефлекс растяжения мышцы и сухожильный рефлекс работают во взаимодействии, обеспечивая базисный механизм быстрого регулирования степени сокращения мышцы. Эти Р. полезны для быстрых приспособлений к изменению положения ноги, когда чел. приходится идти по неровной почве. Разумеется, в локомоции участвуют и др. полисинаптические спинальные Р. Эти Р. включают гораздо больше вставочных нейронов в структуру рефлекторной дуги. Неврологическую основу этих сложных Р. образуют дивергентные (от одного нейрона к неск.) и конвергентные (от неск. нейронов к одному) связи вставочных нейронов.
Моносинаптические и полисинаптические спинномозговые Р. образуют базовый механизм поддержания и приспособления позы. Моторные системы головного мозга влияют на спинномозговые Р. через входные цепи, идущие к вставочным нейронам и γ-мотонейронам. Т. о., изменения спинномозговых Р. могут указывать на патологию в моторных системах головного мозга. Примером этого может служить гиперрефлексия, связанная с травмой латеральных спинномозговых двигательных путей или с повреждением моторных областей лобной доли.
Простейшим рефлексом яв-ся миотатический рефлекс, или рефлекс растяжения мышцы. Этот рефлекс можно вызвать у любой скелетной мышцы, хотя самый известный его пример — коленный рефлекс. Анат. основу миотатического рефлекса составляет моносинаптическая (с одним синапсом) рефлекторная дуга. Она включает сенсорный концевой орган, сенсорное нервное волокно с его клеточным телом в ганглии заднего корешка, α-мотонейрон, на к-ром сенсорный аксон образует синапс, и аксон этого α-мотонейрона, возвращающийся к мышце, от к-рой приходит сенсорное волокно. Сенсорным концевым органом в рефлексе растяжения мышцы служит мышечное веретено. Мышечное веретено имеет мышечные окончания, наз. интрафузальными волокнами, и центральную, немышечную область, связанную с окончанием афферентного нерва. Интрафузальные волокна иннервируются γ-мотонейронами передних корешков спинного мозга. Высшие центры головного мозга могут оказывать влияние на рефлекс растяжения мышцы посредством модуляции активности γ-мотонейронов. Этот рефлекс вызывается растяжением мышцы, что приводит к увеличению длины мышечного веретена и, вследствие этого, к повышению частоты генерирования потенциала действия в сенсорном (афферентном) нервном волокне. Повышенная активность в афферентном волокне усиливает разряд целевого γ-мотонейрона, что вызывает сокращение экстрафузальных волокон мышцы, от к-рой приходит афферентный сигнал. Когда экстрафузальные волокна сокращаются, мышца укорачивается и активность в афферентных волокнах понижается
21.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).
Читайте также:
