
Как называются рефлексы на растяжение мышцы
Мышечные рефлексы (продолжение)
Всем известно, что идеальной техникой бега обладают совсем маленькие дети, которые только что научились ходить. Но как им удается, не имея представления о том, что они делают, выполнять движения с наивысшим КПД. Кроме этого в грудном возрасте все люди крепко хватаются за пальцы - это рефлекс.
Интересно было наблюдать за сыном, когда он еще не умел ходить. Но когда я касался своими большими пальцами его ладошек, он так крепко их обхватывал, что я поднимал его из кроватки. А вот еще один интересный эксперимент, который мы с ним проделывали в то время. Я обхватывал его под мышками и удерживал над диваном так, что он едва касался пальцами ног покрывала. И он начинал выполнять беговые движения. Если при этом я проносил его вперед, то создавалось полное впечатление, что он бежит.
Мышечные рефлексы
В скелетных мышцах имеются два специализированных типа нервных рецепторов, которые могут воспринимать напряжение. Это мышечные веретена и сухожильный орган Гольджи (GTO's). Мышечные веретена имеют сигарообразную форму и состоят из крошечных измененных мышечных волокон, называющихся интрафузалъными волокнами, и нервных окончаний, вместе покрытых соединительнотканной оболочкой. Они располагаются между и параллельно главным мышечным волокнам. Сухожильный орган Гольджи расположен главным образом в месте соединения мышц и их сухожилий или апоневроза.
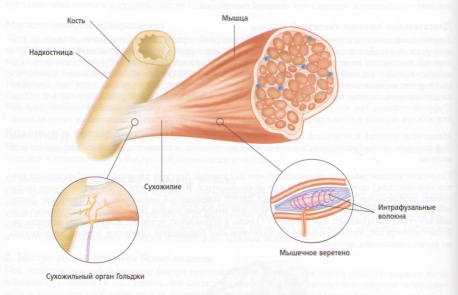
Рис. 18. Анатомия мышечного веретена и сухожильного органа Гольджи
Рефлекс растяжения мышцы
Рефлекс растяжения мышцы обеспечивает сохранение положения тела благодаря поддержанию мышечного тонуса. Он также предотвращает повреждение, давая возможность мышце отвечать на внезапное или неожиданное увеличение длины. Механизм рефлекса следующий:
1. Когда мышца удлиняется, мышечные веретена также растягиваются, заставляя каждое веретено посылать нервный импульс спинному мозгу.
2. При получении импульса спинной мозг немедленно посылает импульс обратно растянутым мышечным волокнам, заставляя их сокращаться, чтобы сопротивляться дальнейшему мышечному растяжению. Этот циркулярный процесс называется рефлекторная дуга.
3. Импульс одновременно посылается от спинного мозга до антагониста мышечного сокращения (т. е. мышце, выступающей против сокращения), вызывая расслабление антагониста, чтобы он не мог сопротивляться сокращению растянутого мышечного волокна. Этот процесс называется реципрокное торможение.
4. Параллельно этому спинальному рефлексу нервные импульсы также посылаются от спинного к головному мозгу с целью передачи информации относительно длины мышцы и скорости ее сокращения. Головной мозг посылает обратно нервные импульсы к мышце для поддержки соответствующего мышечного тонуса, чтобы обеспечить требуемое положение тела и движения.
5. Тем временем чувствительность к растяжению мелких интрафузальных мышечных волокон в пределах мышечного веретена сглаживается и регулируется гамма-эфферентными нервными волокнами, отходящими от двигательных нейронов в спинном мозге. Таким образом, гамма-мотонейрон-рефлекторная дуга обеспечивает равномерность сокращения мышечной ткани, которое иначе было бы хаотичным, если бы мышечный тонус основывался на одном рефлексе растяжения.
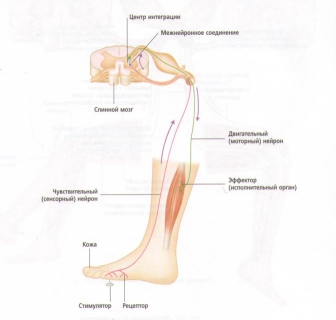
Рис. 19. Основная рефлекторная дуга.
На практике классическим клиническим примером рефлекса растяжения мышцы является коленный рефлекс, или коленный маятникообразный рефлекс, когда по надколенному сухожилию слегка ударяют небольшим резиновым молотком. Рефлекс заключается в следующем:
1. Внезапный удар по надколенному сухожилию приводит к растяжению четырехглавой мышцы.
2. Быстрое растяжение регистрируется мышечными веретенами четырехглавой мышцы, вызывая ее сокращение. Это вызывает небольшой толчок, т. е. внезапное распрямление колена и снятие напряжения мышечных веретен.
3. Одновременно подавляются нервные импульсы к мышцам задней группы бедра, которые являются антагонистами четырехглавой мышцы, это приводит к расслаблению мышц задней группы бедра.
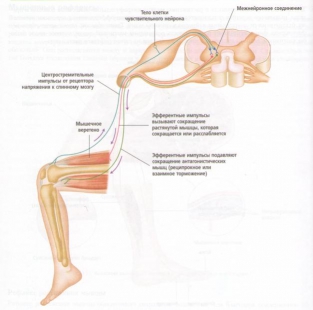
Рис. 20. Рефлекторная дуга напряжения.
На практике другим ярким примером рефлекторной дуги растяжения является следующее: когда человек засыпает в сидячем положении, голова наклоняется вперед, затем отдергивается, потому что напряженные мышечные веретена задней части шеи активизировали рефлекторную дугу.
Рефлекс растяжения мышцы работает постоянно для поддержания тонуса постуральных мышц (т. е. мышц, обеспечивающих правильную осанку). Таким образом рефлекс позволяет человеку оставаться в вертикальном положении без сознательных усилий и без падения вперед. Последовательность событий, предотвращающих падение вперед, происходит за доли секунды следующим образом:
1. В вертикальном положении человек естественно начинает покачиваться вперед.
2. Икроножные мышцы растягиваются, активизируя рефлекс напряжения.
3. Икроножные мышцы последовательно сокращаются, удерживая человека в вертикальном положении.
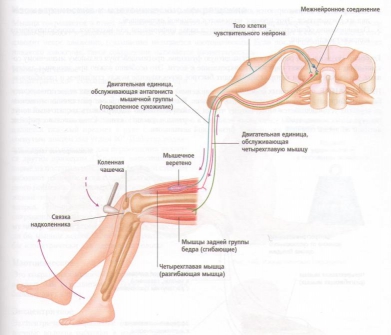
Рис. 21. Коленный маятникообразный рефлекс.
Глубокий сухожильный рефлекс (аутогенное торможение)
В противоположность рефлексу растяжения, который вызывает реакцию мышечного веретена, глубокий сухожильный рефлекс вызывает реакцию сухожильных органов Гольджи, приводящую к мышечному сокращению. Глубокий сухожильный рефлекс вызывает противоположный эффект рефлексу растяжения. Он работает следующим образом:
1. При сокращении мышцы происходит натягивание сухожилий, расположенных с обоих концов мышцы.
2. Напряженность в сухожилии заставляет сухожильный орган Гольджи передавать импульсы спинному мозгу (большое количество импульсов передается мозжечку).
3. Как только импульсы достигают спинного мозга, они подавляют двигательные нервы, обеспечивающие сокращение мышцы, вызывая расслабление.
4. Одновременно активизируются двигательные нервы, обслуживающие мышцы-антагонисты, заставляя их сокращаться. Этот процесс называется взаимной активацией.
5. Одновременно обрабатывается и отправляется назад информация для мозжечка, корректирующего напряженность мышц.
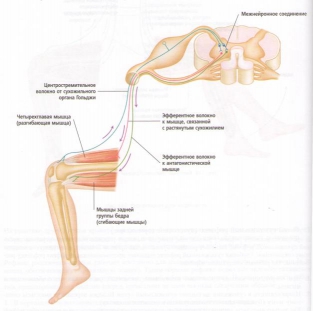
Рис. 22. Глубокий сухожильный рефлекс.
Глубокий сухожильный рефлекс имеет защитную функцию, противодействуя сильному мышечному сокращению, которое может нарушить прикрепление к кости. Это особенно важно при активных движениях, таких, как бег, при которых происходит быстрое переключение между сгибанием и разгибанием.
ПРИМЕЧАНИЕ: при обычных повседневных движениях напряженности в мышцах недостаточно, чтобы активизировать глубокий сухожильный рефлекс. В отличие от этого порог растяжения мышечного веретена устанавливается ниже, потому что он должен постоянно поддерживать достаточный тонус постуральных мышц, чтобы удерживать человека в вертикальном положении. Следовательно, рефлекс активен во время обычной ежедневной физической активности
Нам осталось рассмотреть еще один не маловажный, а даже наоборот - основополагающий вопрос. Это конечно, питание. И связанные с ним мероприятия восстанивления. Не знаю будет ли под силу в одной статье охватить столь жизненный и повседневный вопрос.
Скелетные мышцы состоят из пучков мышечных волокон. Мышечные волокна могут сокращаться, расслабляться или удлиняться.
Мышечные волокна сокращаются под действием электрического импульса, поступающего от нерва. Одно мышечное волокно всегда сокращается полностью, создавая фиксированную силу. Сила, развиваемая всей мышцей, определяется количеством волокон, вовлеченных в это действие нервной системой.
Соединительные ткани в опорно-двигательном аппарате — это связки, сухожилия и фасции. Связки обоими концами крепятся к костям сустава, сухожилия крепят мышцы к костям, а фасции обволакивают группы мышц и пучки мышечных волокон.
Связки и сухожилия содержат большое количество белка коллагена, который придает им жесткость. Связки и сухожилия практически не растягиваются — избыточная нагрузка на эти ткани может привести к травме. Фасции относятся к эластичным соединительным тканям (содержат белок эластин), и их растягивание повышает гибкость. Большинство упражнений на гибкость направлено на растяжение фасций.
При растягивании сначала вытягиваются мышечные волокна, а затем в направлении вытягивающего усилия выравниваются коллагеновые волокна соединительной ткани. Это позволяет упорядочить волокна в направлении растяжения, благодаря чему в упражнениях на растяжение восстанавливается здоровая структура ткани.
При растяжении мышц часть мышечных волокон растягивается, а часть остается в исходном состоянии. Длина мышцы определяется количеством растянутых волокон (аналогично тому, как длина и сила сжатой мышцы определяется количеством сжатых волокон). Чем больше растянутых волокон, тем длиннее растянутая мышца.
В толще мышц находятся нервно-мышечные веретена — нервные окончания, реагирующие на удлинение мышц. Веретена располагаются параллельно мышечным волокнам и растягиваются вместе с ними. При этом они регистрируют как изменение длины мышцы, так и скорость этого изменения.
При сильном или резком удлинении мышцы сигнал от веретен вызывает защитный рефлекс растяжения — мышца самопроизвольно сокращается в попытке помешать удлинению.
Рефлекс растяжения имеет статическую и динамическую составляющие. Статическая составляющая сохраняется все время, пока мышца растянута. Динамическая составляющая — это ответ на скорость изменения длины мышцы, он может быть очень сильным при резком удлинении мышцы и снижается, когда скорость удлинения мышцы уменьшается.
Одна из целей удержания позы растяжения в течение некоторого времени — дать мышечным веретенам адаптироваться к новой длине мышцы с уменьшением интенсивности их сигналов в новом положении.
При сокращении мышца создает натяжение на сухожилии, к которому она крепится. На это реагирует сухожильный орган Гольджи — тип нервного окончания, который находится в местах соединения волокон мышц и сухожилий. Сухожильный орган реагирует на величину возникающей силы натяжения (статический эффект) и на скорость изменения этой силы (динамический эффект: резкое натяжение вызывает сильный сигнал).
Когда сигнал от сухожильного органа превосходит определенный порог, возникает защитная реакция удлинения — резкое самопроизвольное расслабление всей мышцы.
Вторая причина для удержания позы растяжения в течение некоторого времени — позволить произойти реакции удлинения, которая помогает мышце расслабиться (преодолевая рефлекс сжатия).
По отношению к выполняемому движению мышцы можно отнести к одному из четырех типов:
- Агонисты — вызывают движение.
- Антагонисты — вызывают противоположное движение; отвечают за возвращение в начальное положение тела.
- Синергисты — корректируют действие мышцы-агониста для обеспечения нужного направления результирующей силы.
- Стабилизаторы — удерживают остальную часть тела при выполнении движения.
Агонисты и антагонисты обычно находятся с противоположных сторон сустава (например, бицепс и трицепс), а синергисты — на той же стороне, что и агонисты, рядом с ними. При работе крупных мышц-агонистов часто вовлекаются в качестве синергистов находящиеся рядом меньшие мышцы.
Сокращение мышцы-агониста может привести к рефлекторному расслаблению ее антагониста. Это явление называется взаимным торможением.
Взаимное торможение происходит не при всех движениях. Иногда возникает явление совместного сжатия. Например, при приседаниях сжимаются как мышцы брюшного пресса, так и их антагонисты — разгибатели спины.
Упражнения на растяжения легче выполнять с расслабленными, а не с сокращающимися мышцами. Для этого можно использовать эффект взаимного торможения (когда он происходит) — заставляя мышцы-антагонисты расслабляться за счет сжатия агонистов. Также желательно расслабить синергистов мышцы, которую вы растягиваете. Например, при растягивании икроножной мышцы, следует нагнуть стопу, сжав мышцы передней поверхности голени. С другой стороны, икроножная мышца — синергист подколенного сухожилия, поэтому желательно расслабить и его. Для этого нужно выпрямить ногу, сжав тем самым антагониста подколенного сухожилия — четырёхглавую мышцу бедра.

Каждое движение требует координированных действий нескольких мышц: для того, чтобы взять карандаш в руку потребуется участие нескольких мышц, из которых одни должны сокращаться, а другие – расслабляться. Совместно действующие мышцы, т.е. сокращающиеся или расслабляющиеся одновременно называются синергистами, в отличие от противодействующих им мышц-антагонистов. При любом двигательном рефлексе сокращения и расслабления синергисты и антагонисты безупречно координированы друг с другом.
В ответ на растяжение мышцы внешней силой возбуждаются реагирующие только на изменение длины рецепторы мышечных веретён (рецепторы растяжения) (рис. 7.2), которые связаны с особым типом мелких интрафузальных мышечных волокон.

От этих рецепторов возбуждение передаётся по чувствительному нейрону в спинной мозг, где окончание аксона разделяется на несколько ветвей. Одни ветви аксона образуют синапсы с мотонейронами мышц-разгибателей и возбуждают их, что приводит к сокращению мышц: здесь моносинаптический рефлекс – его дуга образована лишь двумя нейронами. Одновременно с этим остальные ветви афферентного аксона активируют деятельность тормозных интернейронов спинного мозга, которые тотчас подавляют активность мотонейронов для мышц-антагонистов, т.е. сгибателей. Таким образом, растяжение мышцы вызывает возбуждение мотонейронов мышц-синергистов и реципрокно тормозит мотонейроны мышц-антагонистов (рис. 7.3).
Силу, с которой мышцы сопротивляются изменению своей длины, можно определить как мышечный тонус. Он позволяет сохранять определённое положение тела (позу). Сила гравитации направлена на растяжение мышц-разгибателей, а их ответное рефлекторное сокращение этому противодействует. Если растяжение разгибателей увеличивается, например, когда на плечи опускается тяжёлый груз, то и сокращение усиливается – мышцы не позволяют себя растягивать и благодаря этому сохраняется поза. При отклонениях тела вперёд, назад или в сторону определённые мышцы растягиваются, а рефлекторное повышение их тонуса сохраняет необходимое положение тела.
По тому же принципу осуществляется рефлекторная регуляция длины у мышц-сгибателей. При любом сгибании руки или ноги поднимается груз, которым может быть и сама рука или нога, но любой груз – это внешняя сила, стремящаяся растянуть мышцы. Ответное сокращение регулируется рефлекторно в зависимости от величины груза.
Сухожильные рефлексы можно вызвать, легко ударив неврологическим молоточком по сухожилию более или менее расслабленной мышцы. От удара по сухожилию такая мышца растягивается и тотчас рефлекторно сокращается.
Последовательность рефлекса: растяжение мышцы вызывает ее сокращение.
Дуга коленного рефлекса (с сухожилия четырёхглавой мышцы бедра):
— внутримышечный рецептор растяжения (в интрафузальном мышечном веретене);
— чувствительный нейрон (тело — в спинномозговом ганглии);
— альфа-мотонейрон (тело — в передних рогах спинного мозга);
— скелетная мышца (четырёхглавой мышцы бедра).
Физиологический смысл рефлекса: поскольку в результате этого рефлекса растяжение (то есть удлинение) мышцы приводит к ее сокращению (то есть укорочению), он направлен на поддержание постоянства длины мышцы. Следовательно, данный рефлекс
— является элементом любых движений, требующих постоянства длины мышц, то есть удержания позы;
— делает движения более плавными, так как препятствует резким изменениям длины мышц.
Эти две функции чрезвычайно важны, и именно поэтому миотатические рефлексы являются самыми распространенными рефлексами спинного мозга.
Рефлексы напряжения
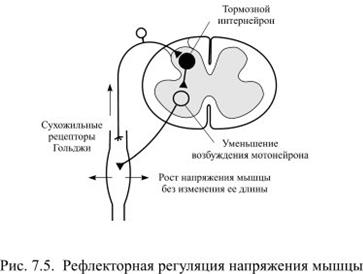
Кроме длины в работающих мышцах рефлекторно регулируется ещё один параметр: напряжение. Когда человек начинает поднимать груз, напряжение в мышцах нарастает до такого значение, чтобы этот груз можно было оторвать от пола, но не более: для подъёма 10 кг не надо напрягать мышцы, как для подъёма 20 кг. Пропорционально росту напряжения усиливается импульсация от проприоцепторов сухожилий, которые называются рецепторами Гольджи (рецепторы напряжения). Это немиелинизированные окончания афферентного нейрона, расположенные между коллагеновыми пучками сухожильных волокон, соединенных с экстрафузальными мышечными волокнами. С ростом напряжения в мышце такие волокна натягиваются и сдавливают рецепторы Гольджи. Нарастающие по частоте импульсы проводятся от них по аксону афферентного нейрона в спинной мозг и передаются тормозному интернейрону, который не даёт мотонейрону возбудиться больше, чем это необходимо (рис. 7.5).
Последовательность рефлекса: напряжение мышцы приводит к ее расслаблению. Дуга рефлекса:
— рецептор напряжения внутри сухожилия (сухожильный орган Гольджи);
— вставочный тормозный нейрон;
Физиологический смысл рефлекса: благодаря данному рефлексу напряжение мышцы приводит к ее расслаблению (растянуть сухожилие и вызвать активацию рецептора можно только при напряжении мышцы). Следовательно, он направлен на поддержание постоянства напряжения мышцы, поэтому:
— является элементом любых движений, требующих постоянства напряжения мышц, то есть удержания позы (например, вертикального положения, требующего достаточно выраженного напряжения мышц-разгибателей);
— препятствует резкому напряжению мышцы, способному привести к травме.
Длина и напряжение мышц взаимообусловлены. Если, например, вытянутая вперёд рука ослабит напряжение мышц, то уменьшится раздражение рецепторов Гольджи, а сила тяжести станет опускать руку. Это приведёт к растяжению мышц, нарастанию возбуждения интрафузальных рецепторов и соответствующей активации мотонейронов. В итоге произойдёт сокращение мышц и рука вернётся в прежнее положение.
Мышечные и сухожильные рецепторы
Какие же рецепторы есть в двигательном аппарате и как они возбуждаются при собственных рефлексах?
Двигательный аппарат отличается наличием очень специализированных и сложно устроенных рецепторных структур, которые могут быть разделены на два основных типа. Рецепторы одного из них (мышечные веретена) расположены в мышцах, а рецепторы другого - в сухожилиях и суставах. Мышечные веретена расположены между мышечными волокнами и одеты капсулой, более широкой в средней части и утончающейся по концам. Внутри каждого веретена также есть интрафузальные мышечные волокна, отличающиеся по строению от обычных (они имеют другое содержание миофибрилл и саркоплазмы и приближаются по строению к эмбриональным волокнам). Важной особенностью мышечных волокон, расположенных внутри веретена, является наличие у них наряду с двигательными окончаниями рецепторных структур. Последние могут быть двух типов - в виде спирали, обвивающей интрафузальное волокно (аннуло-спиральные рецепторы,), и в виде грозди бляшек. Аннуло-спиральные рецепторы соединены с более толстыми афферентными волокнами, а гроздьевидные - с более тонкими.
Двигательные нервные волокна, иннервирующие интрафузальные мышечные волокнам имеют свои особенности. Они значительно тоньше, чем обычные двигательные волокна. Если обычные двигательные волокна относятся в группе А - альфа, по классификации Й..Эрлангера и X.Гассера (1927), то двигательные волокна, иннервирующие мышечные волокна, относятся к группе А -гамма. Они являются аксонами особыхмотонейронов, соответственно также обозначаемых как гамма - мотонейроны.
Рецепторные образования, расположенные в сухожилиях, имеют совершенно другой тип строения. Наиболее характерными здесь являются образования Гольджи. Они состоят из сложной соединительнотканной капсулы, внутрь которой входит афферентное волокно, свивающееся затем в довольно густой клубок. Капсула расположена среди соединительнотканных волокон сухожилия. Инкапсулированные окончания есть также в суставных поверхностях и в фасциях двигательного аппарата.
Кроме этих основных типов, в двигательном аппарате есть и другие, более простые рецепторы, в том числе свободные нервные окончания, Они, по-видимому, не связаны с собственными рефлексами мышц, а представляют собой болевые окончания.
Подробные исследования особенностей афферентного разряда от мышечных рецепторов были проведены в 1933 г. английским физиологом Б.Мэттьюсом, который впервые применил для этой цели отведение потенциалов действия от отдельных афферентных волокон в составе мышечного нерва. Оказалось, что афферентные волокна, идущие от мышцы, по характеру разряда делятся на две основные группы. Если вызвать в мышце одиночное мышечное сокращение, то в одной из этих групп разряд развивается, в основном, в момент укорочения или расслабления мышцы и замолкает в период максимального натяжения, у другой группы, наоборот, разряд усиливается тогда, когда мышца развивает максимальное натяжение.
Несомненно, что эти два типа разрядов соответствуют описанному выше подразделению рецепторов на мышечные веретена и сухожильные рецепторы. Можно себе представить несколько схематически, что механическое действие мышечного сокращения на собственно мышечные и сухожильные рецепторы в какой-то степени противоположно. Сухожильные рецепторы включены как бы последовательно с сократительными элементами, которые механически растягиваются при сокращении. Следовательно, максимальное раздражение таких рецепторов будет наблюдаться как раз на высоте сокращения. Мышечные же веретена расположены по существу параллельно обычным мышечным волокнам. Поэтому при сокращении мышцы интрафузальное волокно окажется, наоборот, расслабленным, и раздражение его рецепторов уменьшится.
Конечно, если мышца растягивается внешней силой, а не сокращается самостоятельно, то особенности раздражения рецепторов изменятся. В этом случае растягиваются как сухожильные, так и интрафузальные структуры, и все типы окончаний будут давать разряд импульсов (хотя порог их раздражения может быть различным).
Специальные исследования зависимости между различными видами собственных рефлексов реакций мышц и особенностями импульсации, поступающей от различных групп рецепторов двигательного аппарата, показали, что основой возникновения как сухожильных рефлексов, так и рефлексов растяжения являются сигналы от рецепторов мышечных веретен. На первый взгляд может показаться странным, почему в таком случае сухожильные рефлексы вызываются воздействием не на мышцы, а на сухожилия, как это подчеркивается и в самом их названии. Однако можно довольно просто убедиться, что к возникновению этих рефлексов рецепторы сухожилия отношения не имеют и что основой рефлекса является вызываемое ударом кратковременное растяжение мышцы. Так, при анестезировании сухожилия удар по нему вызывает такой же сухожильный рефлекс, как и в обычных условиях.
Может показаться странным и то обстоятельство, что возбуждение одних и тех же мышечных рецепторов вызывает две очень различные по своему проявлению рефлекторные реакции. При сухожильном рефлексе это кратковременное фазное движение, а при рефлексе растяжения - длительное тоническое напряжение. Причина различия в этом случае заключается, очевидно, только в том, что в связи с различным видом механического действия афферентная импульсация, поступающая от рецепторов мышечных веретен, оказывается различным образом организованной во времени. Когда наносится удар по сухожилию, то это вызывает кратковременное сильное растяжение и почти синхронный, кратковременный разряд импульсов в соответствующих афферентных волокнах. Если же мышечное веретено растягивается длительно, то в афферентных волокнах возникают не кратковременные синхронные разряды, а постоянная импульсация низкой частоты, длящаяся все время, пока продолжается растяжение мышцы. Постоянная афферентная импульсация и поддерживает тоническое возбуждение соответствующих мотонейронов. Не исключено, конечно, что различные типы окончаний мышечных веретен могут быть в различной степени ответственны за фазный и тонический характер возникающей рефлекторной реакции. Аннуло-спиральные рецепторы сигнализируют в зависимости от скорости растяжения мышцы, они и быстрее адаптируются, гроздьевидные окончания более чувствительны к изменению длины мышцы и медленнее адаптируются, поэтому они дают более стойкий разряд афферентных импульсов, способствуют поддержанию тонического характера рефлекторной реакции.
Если таким образом и сухожильные рефлексы и рефлексы растяжения связаны с возбуждением рецепторов мышечных веретен, то какова же функция рецепторов, расположенных в сухожилиях и, в частности, рецепторов Гольджи? Есть основания считать, что возбуждение рецепторов Гольджи является причиной аутогенного торможения собственных рефлексов мышц. Возбуждение афферентных волокон, которые идут от рецепторов Гольджи, вызывает развитие тормозящих постсинаптических потенциалов в соответствующих мотонейронах и прекращает рефлекторный тонус мышцы при ее чрезмерном растяжении. Известно, что порог возбуждения рецепторов Гольджи при механическом растяжении мышцы выше, чем аналогичный порог возбуждения мышечных веретен. Это различие хорошо согласуется с теми условиями раздражения, при которых развивается аутогенное торможение.
Переходя к рассмотрению центральных механизмов собственных рефлексов мышц, необходимо в первую очередь отметить уже упоминавшуюся характерную особенность сухожильных рефлексов - исключительно короткий скрытый период. Конечно, у рефлексов растяжения измерить скрытый период нельзя, так как это тонический рефлекс, который, постепенно нарастая, длится все время раздражения. Однако можно думать, что поскольку оба рефлекса вызываются возбуждением одних и тех же рецепторов, следовательно, одних н тех же афферентных волокон, то они связаны с деятельностью одних и тех же центральных механизмов.
Одним из условий, которые обеспечивают исключительную быстроту передачи соответствующих нервных влияний, является строение афферентных волокон, которые идут от мышечных рецепторов. Они относятся к самым толстым нервным волокнам (группа А -альфа), имеющим скорость проведения до 120 м/с. Для проприоцептивных афферентных волокон млекопитающих Д.Ллойд и А.Мак-Пита в 1950 г. предложили специальную классификацию, в которой различные группы обозначаются не буквами, а римскими цифрами (группы I, II, III и IV). Группа I соответствует примерно волокнам А - альфа по классификации Й.Эрлангера и X.Гассера и объединяет быстропроводящие волокна, обслуживающие мышечные и сухожильные рецепторы. Афферентные волокна, идущие от сухожильных рецепторов, несколько тоньше по диаметру, чем афферентные волокна, идущие от мышечных рецепторов. Поэтому группа I подразделяется на подгруппу Iа и подгруппу Ib; афферентные волокна от рецепторов мышечных веретен относятся к группе Iа, а афферентные волокна от рецепторов сухожилий - к группе Ib. От части рецепторов мышечных веретен идут и более тонкие волокна, относящиеся к группе II.
Большая скорость осуществления собственных рефлексов мышц обеспечивается, кроме быстроты проведения афферентной волны, также и тем, что рефлекторная дуга таких рефлексов не включает в себя вставочных нейронов.
Трудно сказать, почему в процессе эволюции именно эта рефлекторная дуга оказалась такой быстродействующей. Вероятно, быстрота осуществления собственных рефлексов очень важна для обеспечения нормального функционирования мышечного аппарата и предохранения его от перерастяжения и разрывов при различных двигательных актах.
Сказанное выше касается только возбуждающих эффектов при собственных рефлексах. Центральный скрытый период появления аутогенного торможения всегда оказывается несколько большим, чем центральный скрытый период возбуждающих эффектов (на несколько десятых долей миллисекунд). Это согласуется с общим положением о том, что тормозящие эффекты в центральной нервной системе всегда осуществляются с помощью хотя бы одного дополнительного вставочного нейрона. По-видимому, афферентные волокна, берущие начало от сухожильных рецепторов Гольджи (волокна группы Ib), не контактируют прямо с мотонейронами. Этот путь имеет вставочный нейрон, который превращает возбуждающий эффект в тормозящий. Единственным типом окончаний, которые прямо контактируют с мотонейронами, являются окончания волокон группы Iа от мышечных веретен.
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.
Рефлекса растяжения ( myotatic рефлекс ) является сокращением мышц в ответ на растяжение в мышцах. Это моносинаптический рефлекс , который обеспечивает автоматическое регулирование скелетных мышц длиной.
Когда мышца удлиняется, то мышцы шпиндель растягиваются и его нерва активность возрастает. Это увеличивает нейронную альфа - двигательной активности, в результате чего мышечные волокна сокращаться и , таким образом , сопротивляться растяжению. Вторичный набор нейронов также вызывает противостоящие мышцы для отдыха. Рефлекторные функции для поддержания мышечной массы при постоянной длине.
Гамма - мотонейроны регулируют насколько чувствителен рефлекс растяжения является путем затягивания или расслабления волокон внутри шпинделя. Есть несколько теорий относительно того , что может вызвать гамма - мотонейроны для повышения чувствительности рефлекса в. Например, альфа-гамма - соактивация может держать шпиндели тугой , когда мышца сжимается, сохраняя чувствительность рефлекса растяжения даже мышечные волокна становятся короче. В противном случае шпиндели бы стать слабину и рефлекс перестанет функционировать.
Этот рефлекс имеет самую короткую задержку всех спинальных рефлексов , включая сухожильный рефлекс Гольджи и рефлексы , опосредованный болью и кожные рецепторами.
содержание
- 1 Примеры
- 2 управления супраспинальных
- 3 спинальная контроль
- 4 патологии
- 5 Смотри также
- 6 Ссылки
- 7 Внешние ссылки
Примеры
Человек, стоящий вертикально начинает наклоняться в одну сторону. Постуральная мышца, которые тесно связаны с позвоночником на противоположной стороне будет растягиваться. Мышечные веретена в этих мышцах обнаружат это растяжение, и растянутые мышцы будут сокращаться, чтобы исправить положение.
Другие примеры ( с последующими вовлеченными спинномозговыми нервами ) являются ответами , чтобы растянуть созданный удар при мышечном сухожилии:
Другим примером является группа волокон в икроножных мышц, которые Synapse с моторными нейронами подачи мышечных волокон в одной и той же мышцы. Внезапное простирания, например, нажав ахиллово сухожилие, вызывает рефлекторное сокращение в мышцах , как шпиндели чувствовать растяжку и отправить потенциал действия на двигательные нейроны , которые затем вызывают мышцы сокращаться; этот специфический рефлекс вызывает сокращение в камбаловидной-икроножной группы мышц. Как надколенника рефлекс, этот рефлекс может быть усилен за счет маневра Jendrassik .
супраспинальный контроль
Центральная нервная система может влиять на рефлекс растяжения с помощью гамма-мотонейронов, которые, как описано выше, контроль чувствительности рефлекса.
Ингибирующие сигналы поступают на гамма - нейронах через боковой ретикулоспинальных тракт от Brodmann области 6 , на древней часть мозжечка и красного ядра . Облегчающие сигналы поступают через вентральный ретикулоспинальных тракт от Brodmann области 4 , в neocerebellum и вестибулярного ядра .
Spinal контроль
- Выведение рефлекс
- Демпфирование и загрузка рефлекс
- приём ендрассика
патология
| класс | отклик | Значимость |
|---|---|---|
| 0 | нет ответа | всегда ненормально |
| 1+ | небольшой, но определенно присутствует ответ | может или не может быть нормальным |
| 2+ | бодрый ответ | нормальный |
| 3+ | очень бодрый ответ | может или не может быть нормальным |
| 4+ | подергивание мышц | всегда ненормально |
Ответ Застежка-нож является рефлекс растяжения с быстрым уменьшением сопротивления при попытке согнуть сустав. Это одна из характерных ответных реакций верхнего поражения двигательных нейронов .
Читайте также:
