
Влияние структур головного мозга на активность спинальных рефлексов
Нервная система - сложнейшая и интереснейшая во всем организме. Головной, спинной мозг, а также нервные волокна обеспечивают целостность нашего организма и поддерживают его функционирование. Одна из основных функций нервной системы - защита организма от внешних раздражителей. Это возможно благодаря наличию спинальных рефлексов.

Что такое рефлекс?
Рефлекс - это автоматический ответ организма на внешний раздражитель. Исторически он является одной из наиболее древних реакций нервной системы. Рефлекторный акт - непроизвольный, то есть его невозможно контролировать сознанием.
Последовательность нейронов и их отростков, которые обеспечивают определенный рефлекс, называются рефлекторными дугами. Она необходима для проведения импульса от чувствительного рецептора до нервного окончания в рабочем органе.
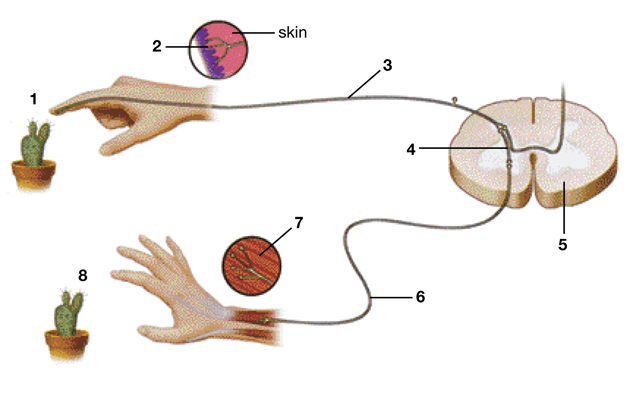
Строение рефлекторной дуги
Рефлекторную дугу двигательного рефлекса называют простейшей, так как она состоит всего лишь из двух нервных клеток или нейронов. Поэтому она также носит название двухнейронной. Проведение импульса обеспечивают следующие отделы рефлекторной дуги:
- Первый нейрон является чувствительным, своим дендритом (коротким отростком) он тянется на периферические ткани, заканчиваясь рецептором. А его длинный отросток (аксон) тянется в другую сторону - к спинному мозгу, заходит в задние рога спинного мозга, а после в передние, образуя соединение (синапс) со следующим нейроном.
- Второй нейрон называется двигательным, его аксон тянется из спинного мозга к скелетным мышцам, обеспечивая их сокращение в ответ на раздражитель. Соединение между нервом и мышечным волокном носит название нервно-мышечного синапса.
Именно благодаря передаче нервного импульса по рефлекторной дуге возможно существование спинальных двигательных рефлексов.
Виды рефлексов
В общем, все рефлексы подразделяют на простые и сложные. Спинальные рефлексы, о которых идет речь в этой статье, относятся к категории простых. Это означает, что для их осуществления достаточно лишь нейронов и спинного мозга. Структуры головного мозга не принимают участия в формировании рефлекса.
Классификация спинальных рефлексов основана на том, какой стимул вызывает данную реакцию, а также в зависимости от функции организма, выполняемой при помощи этого рефлекса. Кроме того, при классификации учитывается, какая часть нервной системы принимает участие в рефлекторном ответе.
Выделяют следующие виды спинальных рефлексов:
- вегетативные - мочеиспускание, потоотделение, сужение и расширение сосудов, дефекация;
- двигательные - сгибательные, разгибательные;
- проприоцептивные - обеспечение ходьбы и поддержание тонуса мышц, возникают при стимуляции мышечных рецепторов.
Двигательные рефлексы: подвиды
В свою очередь, двигательные рефлексы подразделяются на еще два вида:
- Фазные рефлексы обеспечиваются однократным сгибанием или разгибанием мышц.
- Тонические рефлексы возникают при многочисленном последовательном сгибании и разгибании. Они необходимы для поддержания определенной позы.
В неврологии чаще всего применяется другая классификация видов рефлексов. Согласно этому разделению, рефлексы бывают:
- глубокие или проприоцептивные - сухожильные, надкостничные, суставные;
- поверхностные - кожные (проверяются наиболее часто), рефлексы слизистых оболочек.

Методы определения рефлексов
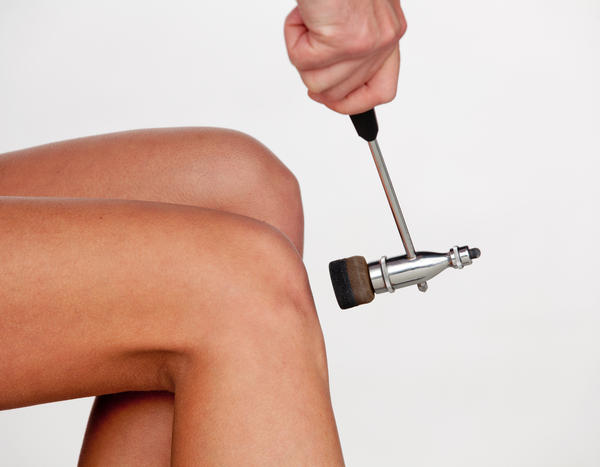
Состояние рефлекса может многое сказать о работе нервной системы. Проверка рефлексов при помощи молоточка - важная часть неврологического осмотра.
Глубокие (проприоцептивные) рефлексы можно определить при помощи легкого постукивания молоточком по сухожилию. В норме должно наблюдаться сокращение соответствующих мышц. Визуально это проявляется разгибанием или сгибанием определенной части конечности.
Кожные рефлексы вызываются быстрым проведением рукояткой неврологического молоточка по специфическим участкам кожи пациента. Эти рефлексы исторически более новые, чем глубокие. Так как они позже образовались, то и при патологии нервной системы именно этот вид рефлексов исчезает первым.
Глубокие рефлексы
Выделяют следующие виды спинальных рефлексов, которые берут свое начало в рецепторе сухожилий:
- Бицепс-рефлекс - возникает при легком ударе по сухожилию двуглавой мышцы плеча, его дуга проходит через IV-VI шейные сегменты спинного мозга (СМ), нормальная реакция - сгибание предплечья.
- Трицепс-рефлекс - происходит при ударе по сухожилию трицепса (трехглавой мышцы), его дуга проходит через VI-VII шейные сегменты СМ, нормальная реакция - разгибание предплечья.
- Пястно-лучевой - вызывается ударом по шиловидному отростку лучевой кости и характеризуется сгибанием кисти, дуга проходит через V-VIII шейные сегменты СМ.
- Коленный - вызывается ударом по сухожилию под надколенником и характеризуется разгибанием ноги. Дуга проходит через II-IV поясничные сегменты спинного мозга.
- Ахиллов - возникает при ударе молоточком по Ахиллову сухожилию, его дуга проходит через I-II крестцовые сегменты спинного мозга, нормальная рефлекторная реакция - подошвенное сгибание стопы.

Кожные рефлексы
Поверхностные, или кожные, рефлексы также важны в неврологической практике. Их механизм подобен глубоким рефлексам: сокращение мышцы, которое возникает при раздражении рецепторных окончаний. Только в данном случае раздражение происходит не при помощи удара молоточка, а штриховым движением рукоятки.
Выделяют следующие виды кожных спинальных рефлексов:
- Брюшные, которые, в свою очередь, подразделяются на верхний, средний и нижний рефлексы. Верхний брюшной рефлекс возникает при раздражении рецепторов участка кожи под реберной дугой, средний - около пупка, нижний - под пупком. Дуги этих рефлексов замыкаются на уровне VIII-IX, X-XI, XI-XII грудных сегментов СМ соответственно.
- Кремастерный - представляет собой подтягивание яичек вверх из-за сокращения его мышц в ответ на раздражение кожного участка внутренней стороны бедра. Дуга рефлекса проходит на уровне I-II поясничных сегментов СМ.
- Подошвенный - сгибание пальцев нижних конечностей при штриховом раздражении кожи подошвы, уровень рефлекса - от V поясничного сегмента до I крестцового.
- Анальный - находится на уровне IV-V крестцовых сегментов и вызывается штриховыми движениями по коже околоанальной области, что приводит к сокращению сфинктера.
Наиболее широкое использование в неврологической практике получило определение брюшных и подошвенного рефлексов.

Патология спинальных рефлексов
В норме рефлексы должны быть оживленными, однофазными (то есть без колебательных движений конечности), умеренной силы. Состояние, когда рефлексы повышенной силы или активности называются гиперрефлексией. Когда же рефлексы, наоборот, снижены, говорят о наличии гипорефлексии. Полное их отсутствие носит название арефлексии.
Гиперрефлексия возникает при повреждении центральной нервной системы. Чаще всего этот патологический симптом возникает при следующих заболеваниях:
- инсульты (ишемический и геморрагический);
- инфекционное воспаление центральной нервной системы (энцефалит, энцефаломиелит);
- церебральный паралич;
- травмы головного и спинного мозга;
- новообразования.
Гипорефлексия, в свою очередь, является одним из проявлений нарушения работы периферической нервной системы. Данное состояние вызывают такие заболевания, как:
- полиомиелит;
- периферические нейропатии (алкогольная, диабетическая).
Однако снижение рефлекторной деятельности нервной системы может возникать и при повреждении центральной нервной системы. Это происходит, когда патологический процесс возникает в том сегменте спинного мозга, где проходит дуга рефлекса. Например, при поражении V шейного сегмента СМ, бицепс-рефлекс будет снижен, в то время как другие глубокие рефлексы, замыкающиеся на более низких сегментах, будут повышены.
Вегетативные рефлексы
Наверное, вегетативные рефлексы являются наиболее сложной разновидностью спинальных рефлексов. Их функцию нельзя определить с помощью обычного неврологического молоточка, однако, именно они обеспечивают жизненно важные функции нашего организма. Их возникновение возможно благодаря функции специфического образования в головном мозге - ретикулярной формации, в которой находятся следующие центры регуляции:
- сосудодвигательный, обеспечивающий деятельность сердца и сосудов;
- дыхательный, который регулирует глубину и частоту дыхания через центры, иннервирующие дыхательные мышцы;
- пищевой, благодаря которому повышается моторная и секреторная функции желудка и кишечника;
- защитные центры, при раздражении которых человек кашляет, чихает, испытывает тошноту и рвоту.
Исследование рефлекторной деятельности нервной системы - важная часть неврологического осмотра пациента, которая позволяет установить локализацию повреждения, что способствует своевременной диагностике.
Общий план организации двигательных систем
Двигательная система – это система, регулирующая работу скелетных мышц, то есть движения. В табл. 1 представлен общий план организации данной системы, а также функции структур этой системы.
Общий план организации двигательной системы
| Структура | Функция, выполняемая изолированной структурой | Роль структуры в осуществлении движения |
| Подкорковые и корковые мотивационные зоны | Побуждение к действию | План |
| Ассоциативные зоны коры | Замысел действия | План |
| Базальные ганглии Мозжечок | Схемы целенаправленных движений (врожденные и приобретенные) | Программа |
| Таламус Двигательная кора | Схемы целенаправленных движений (врожденные и приобретенные) | Программа и ее выполнение |
| Ствол мозга | Регуляция позы | Выполнение |
| Спинномозговые нейроны | Моно- и полисинаптические рефлексы | Выполнение |
| Моторные единицы | Длина и напряжение мышц | Выполнение |
Двигательная система отвечает за осуществление как простых рефлексов (примером может служить отдергивание руки после соприкосновения с горячим предметом), так и сложных поведенческих актов, связанных с процессами высшей нервной деятельности. Из таблицы видно, что двигательная система включает несколько уровней регуляции, низшим из которых является двигательная (моторная) единица. Двигательная единица включает один двигательный нейрон (мотонейрон), который связан с несколькими мышечными волокнами (рис. 1).
В систему регуляции движений вовлечены различные отделы ЦНС, начиная от спинного мозга и заканчивая корой больших полушарий и подкорковыми (базальными) ядрами (ганглиями). При осуществлении двигательного рефлекса импульсы от проприорецепторов (рецепторов мышц и сухожилий) по чувствительному пути поступают в спинной мозг, а от него по восходящим путям направляются в различные отделы головного мозга (рис. 53), и, в конечном счете, в коре больших полушарий осуществляется высший уровень анализа и синтеза поступивших раздражений. Из ЦНС импульсы по двигательным (моторным) путям достигают мышечных волокон, и совершается движение.

Рис. 1. Моторные (двигательные) единицы
Двигательная система выполняет четыре основные функции:
1) отвечает за поддержание определенной позы;
2) способствует ориентации на внешний раздражитель;
3) регулирует перемещение тела в пространстве;
4) обеспечивает манипулирование внешними предметами.
Каждый отдел ЦНС играет определенную роль в регуляции позы и движений.
Роль спинного мозга в регуляции мышечного тонуса и двигательной
Активности
Рефлекторная дуга спинальных рефлексов включает: рецептор, чувствительный (афферентный) путь, чувствительный нейрон (расположен в спинномозговом узле), вставочный нейрон (интернейрон), расположенный в задних рогах спинного мозга, двигательный нейрон (мотонейрон), расположенный в передних рогах спинного мозга, двигательный (эфферентный) путь и эффектор (скелетная мышца, гладкая мышца, железа, сердце). К эффектору импульсы могут идти как по аксонам мотонейронов, так и по постганглионарным волокнам ВНС.
Для поддержания мышечного тонуса достаточно рефлекторной деятельности спинного мозга, в этом случае импульсы к мышце поступают от альфа-мотонейронов спинного мозга.
Двигательные рефлексы, за которые ответственен спинной мозг, осуществляются без участия сознанная, однако связь спинного мозга с головным является необходимой. В процессе регуляции рефлекторной деятельности со стороны отделов головного мозга задействованы гамма-мотонейроны спинного мозга. Так, например, в результате поступления сигналов к гамма-мотонейронам от ретикулярной формации усиливается поток импульсов от проприорецепторов к альфа-мотонейронам спинного мозга.
В спинном мозге расположены:
− центр диафрагмального нерва (3 − 4 шейные сегменты);
− центры мускулатуры верхних конечностей (5 − 8 шейные сегменты);
− центры мускулатуры груди, живота и спины (сегменты грудного отдела);
− центры мускулатуры нижних конечностей (поясничное утолщение);
− вегетативные центры (сегменты грудопоясничного и крестцового отделов).
Выделяют следующие виды рефлексов спинного мозга (спинальных рефлексов):
2. Рефлексы растяжения. В случае быстрого растяжения мышцы, происходит ее сокращение (фазические рефлексы, примером являются сухожильные рефлексы). В случае медленного растяжения мышцы ее длина не изменяется (тонические рефлексы), что необходимо для поддержания равновесия тела в пространстве, так как мышца в этом случае сопротивляется растягивающей ее силе.
3. Рефлексы сгибания и разгибания. Рефлексы сгибания верхних и нижних конечностей наблюдаются после болевого раздражения последних. При этом на противоположной стороне тела часто происходит разгибание конечности. Данное явление связано с тем, что при активации мотонейронов мышц-сгибателей, через контрлатерали аксонов вставочных нейронов на противоположной стороне спинного мозга происходит активация мотонейронов мышц-разгибателей и торможение мотонейронов мышц-сгибателей. В некоторых случаях может иметь место одновременное возбуждение мотонейронов мышц-сгибателей и разгибателей.
4. Ритмические рефлексы. Наблюдаются при раздражении кожи (в случае, например, зуда), когда конечность совершает ритмические чесательные движения; при этом присходит чередование процессов сгибания и разгибание конечности. В данном случае сгибательный и разгибательный центры спинного мозга реципрокно тормозят друг друга. Ритмические рефлексы являются также основой шагательных рефлексов.
5. Рефлекс отталкивания от опоры (при ходьбе, беге).
6. Локомоция. Регуляция спинным мозгом координации движений, что является необходимым для перемещения тела в пространстве. В данном случае спинномозговые рефлекторые центры находятся под контролем ядерных образования ствола мозга, мозжечка, двигательной зоны коры больших полушарий вследствие влияния импульсов, поступающих от экстеро- и проприорецепторов.
При перерезке спинного мозга, в результате чего нарушается связь спинного и головного мозга, такие рефлексы, как защитные, сгибания и разгибания конечностей, сужения сосудов в ослабленном виде сохраняются.
Роль структур головного мозга в регуляции движений и мышечного
Тонуса
Ретикулярная формация среднего мозга может оказывать активирующее влияние на мышечный тонус. Тонус мышц – длительное напряжение мышцы, вызванное регулирующими влияниями нервной системы Под влиянием ретикулярной формации может повышаться чувствительность проприорецепторов, в результате чего усиливается поток импульсов от рецепторов к спинному мозгу и далее к мышце, и мышечный тонус возрастает. Продолговатый мозг, наоборот, оказывает тормозящее влияние на тонус мышц. Причем, действие, как ретикулярной формации, так и ствола мозга в данном случае является неспецифическим, то есть распространяется на различные мышцы. Специфическое влияние различных отделов ЦНС заключается в их действии на отдельные группы мышц. В частности, кора больших полушарий через красные ядра среднего мозга (кортикоруброспинальный тракт), ретикулярную формацию (ретикулоспинальный тракт), а также непосредственно через спинной мозг (корково-спинномозговой или кортикоспинальный тракт) усиливает тонус мышц-сгибателей, а продолговатый мозг посредством вестибулоспинального тракта усиливает тонус мышц-разгибателей.
Ретикулярная формация может оказывать тормозящее и активирующее влияние на осуществление ритмических рефлексов спинного мозга.
Средний и продолговатый мозг, мост играют важную роль в осуществлении статических и статокинетических рефлексов.
Выделяют два типа статических рефлексов: позные (сохраняется поза) и рефлексы выпрямления (например, при переходе из состояния лежа или сидя в состояние стоя). К статическим рефлексам относятся: лабиринтные рефлексы (возникают при изменениях положения головы в пространстве, импульсы поступают от рецепторов вестибулярного анализатора); шейные (возникают при изменениях положения головы относительно туловища, импульсы поступают от проприорецепторов шеи); выпрямительные (импульсы поступают от рецепторов кожи, вестибулярного и зрительного анализаторов). Статокинетические рефлексы помогают сохранить равновесие при действии на организм ускорения (линейный и лифтный рефлексы), а также при повороте головы, туловища в сторону, противоположную движению (вращательный рефлекс).
Голубое пятно, располагающееся на уровне моста, угнетает рефлексы спинного мозга и мышечный тонус в фазу быстрого сна.
Бледное ядро оказывает тормозящее влияние на мышечный тонус, а полосатое тело снимает это влияние.
Средний мозгучаствует в регуляции примитивных движений (без ориентации в пространстве). Верхнее и нижнее двухолмия среднего мозга ответственны за ориентировочные рефлексы на свет и на звук. Красные ядра среднего мозга участвуют в регуляции всех видов движений, так как связаны со спинным мозгом, мозжечком, подкорковыми ядрами, корой больших полушарий. Получая команды от вышележащих центров, красные ядра направляют импульсы по руброспинальному тракту к спинному мозгу и, таким образом, регулируют мышечный тонус. Черная субстанция среднего мозга способствует осуществлению мелких точных действий (например, работа хирурга, часовщика), а также глотательных, жевательных, мимических движений.
Автоматизирование движений и их содружественность находятся под контролем базальных ганглиев (ядер). Бледное ядро оказывает влияние на функционирование заднего и среднего мозга, в частности, как предполагают, оказывает тормозящее влияние на красные ядра среднего мозга. В свою очередь полосатое тело (хвостатое ядро), получая сигналы от таламуса, тормозит рефлекторную деятельность бледного ядра (при недостатке дофамина в хвостатом ядре бледный шар растормаживается, наблюдаются двигательные нарушения), а также оказывает тормозящее влияние на кору больших полушарий. Из хвостатого ядра импульсы поступают в бледный шар и скорлупу, а далее в таламус и чувствительную зону коры больших полушарий, откуда снова направляются к хвостатому ядру. Считается, что базальные ганглии являются высшим эфферентным центром ствола головного мозга и регулируют двигательные безусловные рефлексы, а также вегетативные реакции, сопровождающие данные рефлексы. Предполагается также, что базальные ганглии получают информацию от ассоциативных зон коры больших полушарий, данная информация передается в таламус, куда также поступают сигналы от мозжечка. Собранная информация направляется к двигательной зоне коры больших полушарий, где формируется программа действия (целенаправленного движения), далее импульсы поступают в нижележащие отделы ЦНС, ответственные за выполнение движения.
Мозжечок участвует в поддержание мышечного тонуса: через красные ядра среднего мозга он активирует тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга – тонус мышц-разгибателей. Основную роль в поддержании мышечного тонуса, позы и равновесия тела играет кора червя.
Кора мозжечка(рис. 2)имеет складчатое строение, здесь выделяют доли, каждая из которых, в свою очередь, делится дольки, состоящие из извилин-лепестков (рис. 2, б). В коре выделяют три слоя: молекулярный (наружный), слой клеток Пуркинье и зернистый (внутренний) (рис. 2, в). Афферентные импульсы от проприорецепторов, кожных рецепторов поступают в кору мозжечка по лазающим и моховидным волокнам. По лазающим волокнам импульсы идут от спинного мозга, через оливы продолговатого мозга к клеткам Пуркинье. Моховидные (мшистые) волокна от ядер моста подходят к гранулярным клеткам зернистого слоя (выход импульсов отсюда зависит от деятельности клеток Гольджи), далее направляются к клеткам Пуркинье и в молекулярный слой, где образуют синаптические контакты с корзинчатыми и звездчатыми клетками, аксоны которых направляются к клеткам Пуркинье. Таким образом, последние являются эфферентным выходом коры мозжечка и оказывают тормозящее влияние на ядра мозжечка, которые регулируют активность двигательных центров спинного, продолговатого, среднего и промежуточного мозга.
От подкорковых ядер мозжечка (зубчатого, пробковидного, шаровидного и ядра шатра) (рис. 2, а) импульсы направляются к мотонейронам спинного мозга через ядро Дейтерса продолговатого мозга и ретикулярную формацию. На уровне коры мозжечкаосуществляется программирование движений, их согласование. Роль мозжечка в регуляции двигательной активности заключается в правильном перемещении тела в пространстве, в точном выполнении движений в соответствии с командами, поступающими из коры больших полушарий.

а – мозжечок (вид сзади), ядра мозжечка, расположенные под корой в белом веществе; б – дольки мозжечка; в – строение коры мозжечка; 1 – молекулярный слой, 2 – слой клеток Пуркинье, 3 – зернистый слой
При поражении мозжечка у человека наблюдаются следующие явления:
– атония (от греч. tonos – напряжение) – снижение мышечного тонуса;
– астения (от греч. astheneia – бессилие) – снижение мышечной силы;
– атаксия (от греч. ataxia – беспорядок) – невозможность соотносить выполняемые движения с поставленной целью, нарушение координации и точности движений;
– астазия (от греч. stasis – стояние) – колебательные движения, неспособность стоять;
– дистония – непроизвольное нарушение мышечного тонуса;
– тремор – дрожание частей тела;
– дисметрия – нарушение амплитуды движений (недостаточность или избыточность);
– дизартрия – нарушение моторики речи;
– дизэквилибрия – нарушение равновесия при закрытых глазах.

При участии коры больших полушарий осуществляется высший уровень регуляции мышечного тонуса, важное значение, в данном случае, имеют пирамидные нейроны положения. Большая часть моторной зоны – представительство кистей рук, лица, губ, языка, меньшая часть – представительство туловища и нижних конечностей (рис. 3). Верхние отделы моторной коры отвечают за движение нижних конечностей, а нижние – верхних конечностей. От моторной (двигательной) зоны коры импульсы поступают к базальным ганглиям, стволовой части головного мозга, спинному мозгу и далее к отдельным мышцам.
Рис. 3. Представительство различных
частей тела в прецентральной извилине коры

Взаимосвязь отделов ЦНС, участвующих в осуществлении движений, представлена на рис. 4. Первичная моторная зона коры (4 поле Бродмана, прецентральная извилина) осуществляет непосредственную регуляцию тонуса двигательных центров ствола головного мозга и спинного мозга на основе информации, поступившей из соматосенсорной зоны. Во вторичную моторную кору (6 поле Бродмана) импульсы приходят из ассоциативных зон коры; на основе поступившей информации во вторичной коре формируется программа выполнения движений, которая направляется в первичную кору, то есть первичная кора подчиняется вторичной. От премоторной зоны импульсы направляются к базальным ганглиям, к красным ядрам и черной субстанции среднего мозга, к продолговатому мозгу и ретикулярной формации и далее по рубро-, ретикуло- и вестибулоспинальным трактам – к спинному мозгу.
Рис. 4. Взаимосвязь отделов ЦНС, участвующих
в осуществлении движений (http://www.ido.rudn.ru)
Посредством данных импульсов регулируется двигательная активность скелетных мышц, дыхательной мускулатуры, плавность ритмических движений, осуществляется объединение отдельных движений в двигательный акт.
Нижнетеменные области коры способствуют точной адресации команд к определенным мышцам. Переднелобные зоны участвуют в программировании и регуляции произвольных движений. Зоны коры, относящиеся к лимбической системе, обеспечивают эмоциональную окраску движений.
Функциональной единицей нервной деятельности является рефлекс как ответная реакция нервной системы на раздражение.
Оценка сухожильных и кожных безусловных рефлексов широко используется в клинике нервных болезней для определения очага поражения, его уровня, т.е. для установления топического диагноза. А в сочетании с другими неврологическими симптомами это дает возможность поставить и клинический диагноз. Например, острое нарушение мозгового кровообращения, опухоль головного и спинного мозга, травматическое повреждение ЦНС.
Соматические спинномозговые рефлексы человека Таблица 1
Сухожильные проприорецептивные рефлексы:
При вызывании рефлексов необходимо добиваться от больного возможно более полного расслабления исследуемой конечности. Удары молоточком следует наносить с одинаковой силой. При оценке рефлексов обращают внимание на их выраженность и симметричность. Поэтому исследовать одни и те же рефлексы надо справа и слева и сразу их сравнивать. Выраженность рефлексов у здоровых людей может индивидуально колебаться, в частности, симметричное положение или оживление рефлексов при полной сохранности мышечной силы можно расценивать как вариант нормы.
Наблюдаются следующие изменения рефлексов: понижение или утрата (при повреждении рефлекторной дуги), повышение, и извращение (при поражении пирамидной системы и растормаживании сегментарного аппарата спинного мозга).
СПИНАЛЬНЫЙ ШОК
Спинальный шок - это обратимое угнетение двигательных и вегетативных рефлексов, возникающее после перерыва спинного мозга.
Существуют два взгляда на природу спинального шока:
1 .Спинальный шок является следствием выпадения возбуждающего влияния вышележащих отделов ЦНС.
2.Спинальный шок возникает в результате устранения подавляющего влияния высших моторных центров на спинальное торможение.
Наиболее вероятно, по-видимому, существование обоих путей.
Симптомы повреждения спинного мозга складываются из нарушения основных функций спинного мозга (сегментарной рефлекторной деятельности и проводниковой функции).
Сегментарные нарушения возникают при поражении серого вещества задних (чувствительных) и передних (двигательных) корешков. Они проявляются: двигательными, чувствительными, рефлекторными, сосудистыми, секреторными, трофическими расстройствами.
При поражении задних корешков развивается утрата всех видов чувствительности в соответствующей дерматоме, возможно ослабление или утрата рефлекторной дуги, которое проходит через данный корешок. При поражении передних корешков развивается парез (ослабление) двигательной функции или вялый паралич (отсутствие двигательной функции с расслаблением скелетной мускулатуры и ее последующей атрофией) мышц, соответствующих миотомов. При сегментарном поражении клеток боковых рогов спинного мозга возникают вегетативные, сосудистые,
трофические расстройства, нарушения потоотделения, пилоромоторной реакции, рефлекторных функций тазовых органов.
Проводниковые нарушения возникают при поражении проводящих путей спинного мозга.При поражении двигательных пирамидных путей в состоянии спастического паралича (отсутствие движений с явлениями повышения мышечного тонуса) оказываются все мышцы', иннервируемые от сегментов, расположенных ниже поражения спинного мозга. При перерыве чувствительных путей наступает анестезия (потеря чувствительности) книзу от уровня, охватывающая зоны иннервации всех расположенных ниже сегментов.
ОСНОВНЫЕ ПРОЯВЛЕНИЯ ПОРАЖЕНИЯ СПИННОГО МОЗГА НА РАЗНЫХ УРОВНЯХ
1 .Полный перерыв спинного мозга в верхнешейном отделе (С1-С4):
- вялый паралич мышц шеи,
- спастическая тетраплегия (отсутствие двигательной активности всех конечностей с повышением тонуса мышц),
- нарушение функции тазовых органов (нарушение моче- и калоотделения).
2.Полный перерыв на уровне шейного утолщения (C5 -т1):
- вялый паралич верхних конечностей с атрофией мышц,
- спастический паралич ног с арефлексией,
- общая анестезия ниже уровня поражения,
- нарушение функции тазовых органов по центральному типу (задержка и периодическое недержание мочи и кала).
3.Полный перерыв на уровне грудных сегментов:
- нижняя спастическая параплегия,
- общая анестезия нижней части тела,
- нарушение функции тазовых органов (задержка моче- и калоотделения),
- при нарушении верхних и средних грудных сегментов (Т2 –Т6) - нарушение функции межреберных мышц (паралич-дыхания),
- при разрушении Т10 –Т12- паралич мышц брюшного пресса.
4.Полный перерыв на уровне пояснично-кресцового утолщения (L1-S2):
- вялый паралич нижних конечностей с атрофией мышц,
- анестезия нижний конечностей,
- нарушение потоотделения и пилоромоторной реакции на ногах,
- нарушение функции тазовых органов.
5.Поражение сегментов S3-Со1
- нарушение функции тазовых органов по периферическому типу (истинное недержание мочи и кала, отсутствие позывов),
- анестезия аногенитальной зоны,
- параличи и боли отсутствуют.
Полная параплегия, возникающая при разрывах спинного мозга, сопровождается выключением всех видов произвольных движений, утратой чувствительности, а также временной арефлексией (исчезновением рефлекторной дуги), которые проходят каудальнее места перерыва спинного мозга.
ПРИ ПОРАЖЕНИИ ТОЛЬКО ЧАСТИ СПИННОГО МОЗГА картина складывается из различных сочетаний нарушений движения, координации, чувствительности, расстройств функции тазовых органов, трофики пораженной зоны (пролежни) и т.д.
1 .Поражение передней половины поперечника спинного мозга:
- периферические параличи мышц соответствующего миотома,
- центральный паралич мышц и анестезия ниже очага поражения,
- нарушение функции тазовых органов по центральному типу (нет произвольной координации процессов).
2.Поражение одной половины спинного мозга:
- снижение мышечно-суставной чувствительности на стороне поражения,
- проводниковая потеря поверхностной чувствительности на противоположенной стороне.
3.Поражение задней трети спинного мозга:
- проявляется нарушением глубокой чувствительности, сенситивной атаксией.
В последующие недели и месяцы двигательные рефлексы восстанавливаются. СТАДИИ ВОССТАНОВЛЕНИЯ:
1) полная арефлексия (4-6 недель);
2) восстановление небольших рефлекторных движений пальцев (после 2-х недель - до нескольких месяцев);
3) стадия постепенного усиления сгибательных рефлексов. Генерализованные сгибательные рефлексы постепенно начинают сопровождаться перекрестными разгибательными. При этом важнейшей рефлексогенной зоной является стопа;
4) хроническая стадия (спустя 6 и более месяцев после травмы) - характеризуется преобладаем генерализованных сгибательных рефлексов, могут появляться и разгибательные рефлексы, способные переходить в длительные разгибательные спазмы, обеспечивающие даже так называемое "спинальное состояние" - больные могут стоять некоторое время без поддержки.
Эти стадии используются для прогноза заболевания, восстановления некоторой двигательной активности пациентов путем тренировки и протезирования.
ВЕГЕТАТИВНЫЕ РЕФЛЕКСЫ СПИННОГО МОЗГА И ДИНАМИКА ПОСЛЕ ТРАВМЫ
Ниже места повреждения спинного мозга вегетативные рефлексы (сосудо-двигательные. потоотделительные, рефлексы опорожнения мочевого пузыря и прямой кишки, половые) исчезают на несколько месяцев, а произвольная их регуляция при полном перерыве спинного мозга не восстанавливается.
В течение 1-2 месяцев кожа соответствующих областей сухая и покрасневшая вследствие снижения симпатического тонуса. Через несколько месяцев эта реакция сменяется явлениями гиперрефлексии: усиление сосудосуживающих и потоотделительных рефлексов в ответ на раздражение кожи, сокращение мышц сгибателей, растяжение прямой кишки, сокращение мочевого пузыря.
Рефлексы эвакуации мочевого пузыря восстанавливаются лишь спустя месяцы и возникают лишь в ответ на раздражение соответствующих областей (поколачивание по нижней части передней брюшной стенки - опорожнение мочевого пузыря, растяжение анального отверстия - опорожнение прямой кишки).
Таким образом, изолированный спинной мозг человека может выполнять основные регуляторные вегетативные функции, что играет важную роль при соответствующей тренировке и правильном лечении в обеспечении вегетативных функций у больных с пара- и тетраплегиями.
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ
Ретикулярная формация мозгового ствола была впервые описана русским ученым В.М. Бехтеревым (1898) как диффузное скопление нейронов, простирающихся через весь ствол головного мозга от верхних шейных спинальных сегментов до промежуточного мозга. Анатомически она может быть разделена на ретикулярную формацию продолго-ватого мозга, варолиевого моста и среднего мозга. Вместе с тем, в функциональном отношении в ретикулярной формации разных отделов мозгового ствола есть много общего. Поэтому целесообразно рассматривать ее как единую структуру.
Для ретикулярной формации характерным является то, что её многочисленные нейроны образуют как бы диффузную сеть (лат. reticulae - сеть, отсюда и название этой структуры головного мозга), которая пронизана многочисленными нервными волокнами. Часть из них имеет нисходящее направление к нейронам спинного мозга и образует ретикулоспинальные пути, а часть - восходящее к коре больших полушарий, например, сенсорные восходящие системы. Обилие волокон поступает к нейронам ретикулярной формации из мозжечка, таламуса, базальных ядер, коры больших полушарий. Таким образом, по организации своих афферентных и эфферентных связей эта система приспособлена к объединению влияний из различных мозговых структур, а выходящие из нее пути оказывают в свою очередь влияния как на вышележащие, так и на нижележащие мозговые центры.
При микроэлектродном исследовании нейронов ретикулярной формации выяснилось, что близкорасположенные клетки могут обладать совершенно различными функциональными характеристиками. Следовательно, организация межнейронных связей в ретикулярной формации достаточно дифференцирована и отдельные ее клетки соединены между собой довольно специфичными связями. В настоящее время в составе ретикулярной формации мозга человека описано более 100 ядер.
Неспецифические нисходящие влияния. В 1946 г. американский нейрофизиолог Х.Мегоун и его сотрудники обнаружили, что ретикулярная формация мозгового ствола имеет непосредственное отношение к регуляции не только вегетативной, но и соматической рефлекторной деятельности: ретикулярная формация изменяет течение спинальных двигательных рефлексов. Пути, тормозящие а- и у-мотонейроны сгибателей и активирующие разгибателей, начинаются в области варолиева моста, в то время как пути от бульбарного отдела оказывают противоположное действие.
Первые работы Х.Мегоуна показали, что раздражение гигантоклеточного ядра ретикулярной формации вызывает ослабление в одинаковой мере всех спинальных двигательных рефлексов и сгибательных, и разгибательных, т.е. нисходящая система, берущая начало в вентрокаудальной части ретикулярной формации, имеет неспецифическую тормозящую функцию. Раздражение более дорсальных и оральных ее участков вызывает, наоборот, диффузное облегчающее действие на спинальную рефлекторную деятельность.
Восходящие влияния. Они связаны с регуляцией активности высших отделов мозга, главным образом, коры больших полушарий. Данные об этом были получены еще в тридцатые годы прошлого столетия бельгийским нейрофизиологом Ф.Бремером (1935). Он, проводя перерезку головного мозга на различных уровнях, сделал вывод, что для бодрствующего состояния высших отделов мозга необходимо непрерывное поступление к ним афферентной импульсации. В состав ретикулярной формации мозгового ствола входят структуры, при возбуждении которых животное просыпается и
становится активным (настораживается, принюхивается), но и структуры, активация которых вызывает засыпание животного. Это ядра шва. В окончаниях аксонов нейроны этих ядер выделяют серотонин. В опытах на животных было показано, что локальное разрушение этих ядер приводит к хронической бессонице, которая заканчивается смертью. Известно, что истощение серотонина мозга у человека также приводит к бессонице.
Ретикулярная формация- продолговатого мозга включает три поля: гигантоклеточное, крупноклеточное и мелкоклеточное, два ядра - латеральное ретикулярное и парамедианное и несколько групп катехоламиновых нейронов. Гигантоклеточное поле ретикулярной формации является важным звеном передачи в центр таламуса болевой информации, поступающей из спинного мозга. Здесь обнаружено большое количество опиатных рецепторов и эндогенных пептидов,
участвующих в антиболевом механизме. Крупноклеточное поле тесно связано со спинным мозгом, среднемозговой ретикулярной формацией и парафасцикулярным ядром таламуса. Аксоны ретикулярных нейронов крупноклеточного поля скапливаются в ретикулярной формации варолиева моста. Катехоламиновые группы нейронов входят в ядро одиночного тракта. Они принимают афферентацию от барорецепторов сосудов и передают информацию в гипоталамус и латеральную тегментальную группу.
Ретикулярная формация моста включает в себя группу ядер и полей, имеющих структурные, химические и функциональные различия: гигантоклеточное поле, тегментальное ретикулярное ядро Бехтерева, паралемнисковое поле, голубое пятно, парабрахиальную область. Последняя содержит разнородные группировки нейронов, имеющих отношение к вкусовой, двигательной и зрительной
функциям и посылающих прямые проекции в кору.
Голубое пятно является наиболее крупным объединением норадренергических нейронов, небольшую часть составляют серотонинергические клетки. Нейронные группировки голубого пятна связаны с реакцией самораздражения, циклом сна-бодрствования, регуляцией сосудистого тонуса, восприятием боли. Паралемнисковое поле состоит из орального и каудального ядер моста, активность нейронов связана с движением глаз в быстроволновой фазе сна.
Ретикулярная формация среднего мозга занимает обширную часть покрышки мозгового ствола и содержит несколько разнородных нейронных объединений, включенных в разные функциональные подсистемы. К ним относят центральное поле покрышки, латеральный отдел покрышки, кунеинформное ядро, вентральное поле покрышки. Центральное поле покрышки получает афферентные проекции от нижележащих образований, ядер мозжечка, центрального серого вещества,
латерального гипоталамуса, моторной и сенсорной коры. Его выходы поступают в вентральные и медиальные ядра таламуса, верхние бугры четверохолмия, гипоталамус, нижележащие отделы мозгового ствола.
Латеральный отдел покрышки имеет тесные связи с вентромедиальным ядром гипоталамуса, наружным коленчатым телом.
Вентральное поле покрышки обеспечивает дофаминергическую иннервацию структур гипоталамуса, подкорковых ядер и новой коры, ее рассматривают как ретикулярный компонент системы черная субстанция - стриопаллидарная система. Результаты раздражения, приводящие к значительному усилению двигательной активности, эмоциональности, самостимуляции, свидетельствуют об участии вентрального поля покрышки в процессах висцеромоторной интеграции.
Кунеинформное ядро (клинообразное) имеет обширные связи как со слуховой, зрительной, двигательной и другими специализированными системами, так и теснейшие связи с ретикулярной формацией всех уровней.
ТОНИЧЕСКИЕ РЕФЛЕКСЫ
Существует 2 типа соматических рефлексов - фазные и тонические. Фазные рефлексы являются основой сложных координированных локомоторных актов. Они обеспечивают перемещение тела в пространстве (ходьбу, плавание, бег) или его частей в пространстве. Тонические рефлексы направлены на сохранение естественной позы, т.е. определённой ориентации тела в пространстве, определённого взаимного расположения его частей (у человека - выпрямление позвоночника, стояние на двух ногах, вертикальное положение головы).
Тонические рефлексы в свою очередь подразделяются на статические и статокинетические: статические рефлексы возникают при изменениях положения тела; статокинетические рефлексы при перемещении тела в пространстве или как реакция на ускорение.
1. Статические рефлексы
Статические рефлексы разделяются на рефлексы позы (позно-тонические), которые обеспечивают сохранение естественного положения тела, и на выпрямительные рефлексы, которые восстанавливают нормальную позу.
Позно-тонические рефлексы
Позно-тонические рефлексы возникают при изменениях положениях головы по отношению к туловищу. В этих условиях в связи с перемещением центра тяжести тела появляется опасность нарушения равновесия. Одновременно раздражаются отолитовый аппарат вестибулярного анализатора, проприорецепторы мышц и сухожилий, что и дает начало позно-тоническим рефлексам. Эти рефлексы приводят к перераспределению тонуса мышц шеи, туловища и конечностей, что обеспечивает поддержку ток части тела, куда сместился центр тяжести.При активных поворотах головы перераспределение мышечного тонуса создает базу для последующего перемещения всего тела. Поэтому у хорошо тренированного спортсмена поворот головы всегда предшествует повороту туловища.
В связи с тем, что основным источником афферентной импульсации для позно-тонических рефлексов являются проприорецепторы мышц шеи и рецепторы вестибулярного анализатора, рефлексы первого типа называются шейными, а второго — вестибулярными (лабиринтными).
Шейные тонические рефлексы
Шейные тонические рефлексы запускаются при возбуждении проприоцепторов мышц шеи. Как показали опыты Р. Магнуса, шейные тонические рефлексы в зависимости от положения головы относительно туловища вызывают следующие изменения тонуса у животных:
1 .откидывание головы назад - возрастание тонуса разгибателей передних и уменьшение тонуса задних конечностей;
2.опускание головы - понижение раз-
гибательного тонуса передних конечностей
при увеличении тонуса разгибателей задних
конечностей;

3.наклон или поворот головы - компенсаторное усиление разгибательного тонуса передней и задней конечностей на той половине тела, в сторону которой произошел наклон (или поворот) головы. Это так называемый рефлекс вращения по Магнусу.
В отличие от этого, у человека наклон головы вперед повышает тонус всех мышц сгибателей, а назад — разгибателей. Наклон головы к плечу приводит к повышению тонуса мышцсгибателей этой стороны и разгибателей — противоположной.
Все шейные рефлексы являются полисинаптическими. Импульсы от проприоцепторов шейных мышц идут в соответствующие центры продолговатого мозга, а оттуда нисходящие команды поступают к мотонейронам спинного мозга и оказывают определенное влияние на позный тонус.
Дата добавления: 2019-02-12 ; просмотров: 1433 ;
Читайте также:
