
Корково спинальный пирамидный путь
Существуют следующие нисходящие проводящие пути:
• корково-спинномозговой проводящий путь (пирамидный проводящий путь);
• ретикуло-спинномозговой проводящий путь (экстрапира-мидный путь);
• преддверно-спинномозговой проводящий путь;
• покрышечно-спинномозговой проводящий путь;
• шовно-спинномозговой проводящий путь;
• проводящие пути аминергических систем ЦНС;
• проводящие пути вегетативной нервной системы.
Корково-спинномозговой проводящий путь
Корково-спинномозговой проводящий путь представляет собой крупный проводящий путь произвольной двигательной активности. Около 40 % его волокон начинается из первичной моторной коры прецентральной извилины. Остальные волокна берут начало из дополнительной моторной области на медиальной стороне полушария, премоторной коры головного мозга на латеральной стороне полушария, соматической сенсорной коры, коры теменной доли и коры поясной извилины. Волокна от двух вышеупомянутых сенсорных центров заканчиваются на чувствительных ядрах ствола головного мозга и спинного мозга, где они регулируют передачу чувствительных импульсов.
Корково-спинномозговой проводящий путь спускается вниз через лучистый венец и заднюю ножку внутренней капсулы к стволу головного мозга. Затем он проходит в ножке (головного мозга) на уровне среднего мозга и базилярной части моста, достигая продолговатого мозга. Здесь он образует пирамиду (отсюда название — пирамидный проводящий путь).
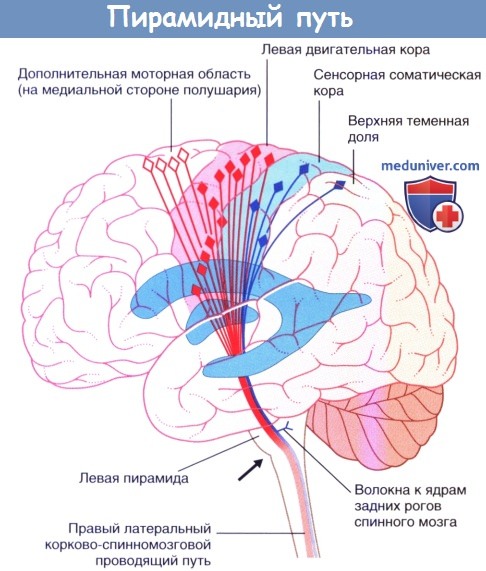
Демонстрация хода волокон пирамидного пути с левой стороны.
Дополнительная моторная область на медиальной стороне полушария.
Стрелкой показан уровень перекреста пирамид. Чувствительные нейроны выделены синим цветом. 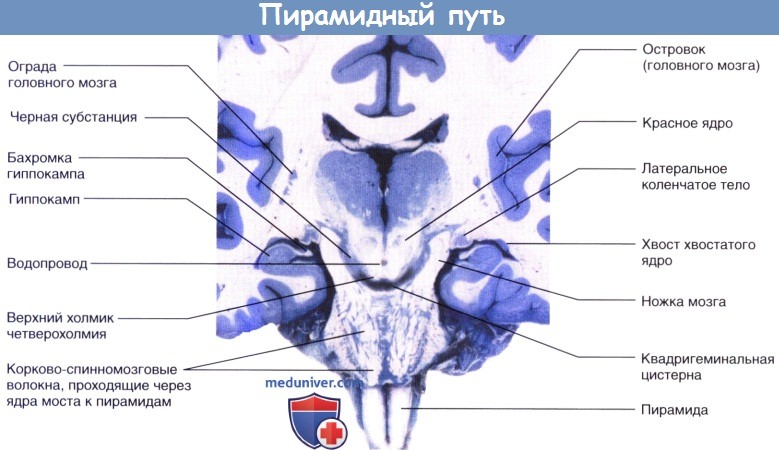
Коронарный срез бальзамированного головного мозга пациента с последующей обработкой сульфатом меди (окраска по Маллигану),
демонстрирующий неокрашенные корково-спинномозговые волокна, идущие через ядра моста в сторону пирамид.
Характеристика волокон корково-спинномозгового пути выше уровня спинномозгового перехода:
• около 80 % (70-90 %) волокон переходят на противоположную сторону на уровне перекреста пирамид;
• эти волокна спускаются по противоположной стороне спинного мозга и составляют латеральный корково-спинномозговой проводящий путь (перекрещивающийся корково-спинномозговой проводящий путь); оставшиеся 20 % волокон не перекрещиваются и продолжают спускаться вниз в передней части спинного мозга;
• половина из этих неперекрещивающихся волокон вступает в передний/вентральный корково-спинномозговой проводящий путь и располагается в вентральном/переднем канатике спинного мозга на шейном и верхнем грудном уровнях; данные волокна переходят на противоположную сторону на уровне белой спайки и иннервируют мышцы передней и задней стенок брюшной полости;
• другая половина вступает в латеральный корково-спинномозговой проводящий путь на своей половине спинного мозга.
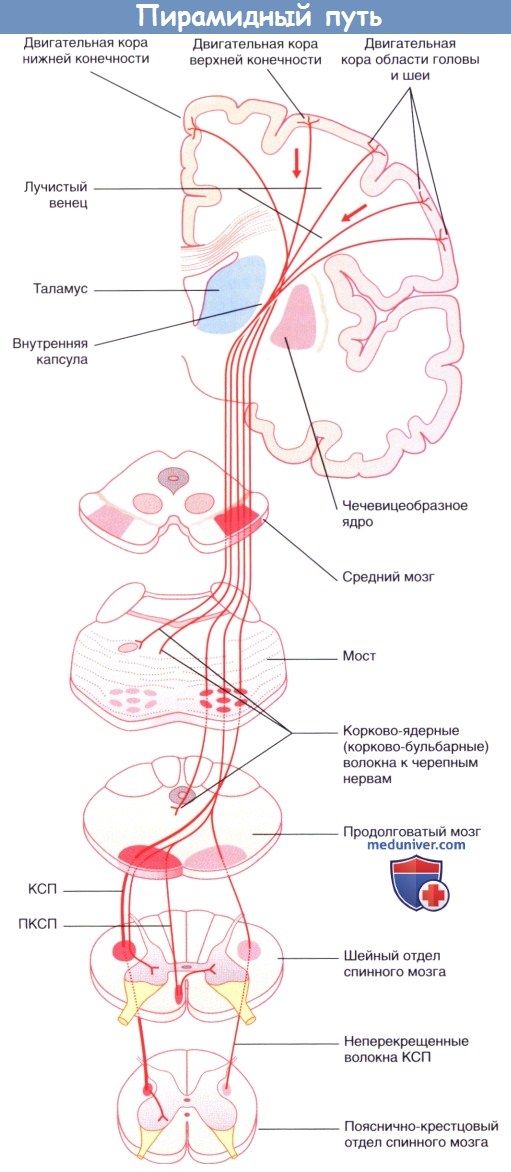
Пирамидный проводящий путь.
КСП — корково-спинномозговой проводящий путь;
ПКСТ — передний корково-спинномозговой проводящий путь;
ЛКСП — латеральный корково-спинномозговой проводящий путь.
Обратите внимание: показан только двигательный компонент; компоненты теменной доли опущены.
Клетки-мишени латерального корково-спинномозгового пути:
Двигательная единица — это комплекс, состоящий из нейрона переднего рога спинного мозга и всех мышечных волокон, которые этот нейрон иннервирует. Нейроны малых двигательных единиц избирательно иннервируют небольшое количество мышечных волокон и участвуют в выполнении тонких и точных движений (например, при игре на пианино). Нейроны переднего рога, иннервирующие крупные мышцы (например, большую ягодичную мышцу), способны по отдельности вызвать сокращение сотни мышечных клеток сразу, так эти мышцы отвечают за грубые и простые движения.
Фракционирование имеет большое значение при выполнении привычных движений, таких как застегивание пальто или завязывание шнурков. Травматическое или другое повреждение корковомотонейронной системы на любом уровне влечет за собой утрату навыков выполнения привычных движений, которые затем редко поддаются восстановлению.
При выполнении данных движений α- и γ-мотонейроны активируются совместно через латеральный корково-спинномозговой проводящий путь таким образом, что веретена мышц, первично задействованных в движении, посылают импульсы об активном растяжении, а веретена мышц-антагонистов — о пассивном растяжении.
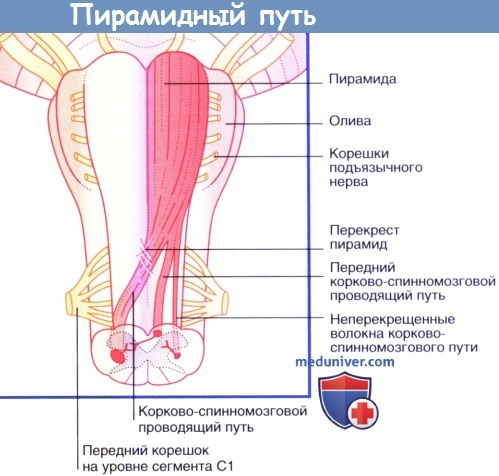
Продолговатый мозг и верхние отделы спинного мозга, вид спереди.
Продемонстрированы три группы нервных волокон левой пирамиды.
б) Клетки Реншоу. Функции синапсов латерального корково-спинномозгового пути на клетках Реншоу довольно многочисленны, так как торможение на некоторых клеточных синапсах главным образом происходит за счет интернейронов типа Iа; на других синапсах данную функцию выполняют клетки Реншоу. Вероятно, наиболее важная функция — контроль совместного сокращения основных движущих мышц и их антагонистов для фиксации одного или нескольких суставов, например при работе с кухонным ножом или лопатой. Совместное сокращение происходит за счет инактивации ингибирующих интернейронов Iа клетками Реншоу.
в) Возбуждающие интернейроны. Латеральный корково-спинно-мозговой проводящий путь влияет на деятельность двигательных нейронов, расположенных в средней части серого вещества и в основании переднего рога спинного мозга, иннервирующих осевые (позвоночные) мышцы и мышцы проксимальных отделов конечностей посредством возбуждающих интернейронов. г) la-ингибирующие интернейроны. Эти нейроны также расположены в средней части серого вещества спинного мозга и активируются латеральным корково-спинномозговым путем в первую очередь при совершении произвольных движений.
Активность Ia-интернейронов способствует расслаблению мышц-антагонистов до того, как начнут сокращаться мышцы-агонисты. Кроме того, они вызывают рефрактерность мотонейронов мышц-антагонистов к стимуляции афферентами нервно-мышечного веретена при их пассивном растяжении во время движения. Последовательность процессов при произвольном сгибания коленного сустава показана на рисунке ниже.
С другой стороны, во время произвольного сгибания коленного сустава мышцы способствуют данному движению с помощью такого же механизма, но уже через рефлекс помощи. Изменение знака с отрицательного на положительный называют рефлексом перемены направления.)
д) Пресинаптические ингибиторные нейроны, обеспечивающие рефлекс растяжения. Рассмотрим движения спринтера. На каждом шаге сила тяжести тянет его тело вниз, на выпрямленное четырехглавой мышцей колено. В момент соприкосновения с землей все нервно-мышечные веретена в сокращенной четырехглавой мышце резко растягиваются, в результате чего возникает опасность разрыва мышцы. Сухожильный орган Гольджи обеспечивает некоторую защиту посредством внутреннего торможения, однако основной защитный механизм обеспечивает латеральный корково-спинномозговой путь через пресинаптическое торможение афферентов веретен вблизи их контакта с мотонейронами.
В то же время удлинение паузы до ахиллового рефлекса служит преимуществом в этой ситуации, так как происходит восстановление мотонейронов, иннервирующих заднюю часть голени, для следующего рывка. Предполагают, что степень подавления рефлекса растяжения со стороны латерального корково-спинномозгового пути зависит от конкретных движений.
е) Пресинаптическое ингибирование чувствительных нейронов первого порядка. В заднем роге серого вещества спинного мозга существует некоторое подавление передачи чувствительных импульсов в спиноталамический проводящий путь при совершении произвольных движений. Это происходит путем активации синапсов, образованных ингибирующими вставочными нейронами и первичными чувствительными нервными окончаниями.
Еще более тонкую регуляцию наблюдают на уровне тонкого и клиновидного ядер, где волокна пирамидного пути (после пересечения) способны усиливать передачу чувствительных импульсов во время медленных аккуратных движений или ослаблять ее во время совершения быстрых движений.
Видео урок анатомия пирамидных путей - tractus corticospinalis et corticonuclearis
Редактор: Искандер Милевски. Дата публикации: 15.11.2018
Эфферентные проводящие пути классифицируют на пирамидные и экстрапирамидные. Низшие позвоночные пирамидных путей не имеют – они появляются только у млекопитающих и достигают наибольшего развития у человека.
Пирамидные пути начинаются в пятом слое коры от гигантских пирамидных клеток (клеток Беца), находящихся в предцентральной извилине и парацентральной дольке. Заканчиваются эти тракты на нейронах двигательных ядер черепных нервов и двигательных ядер передних рогов спинного мозга. В стволе головного мозга пирамидные пути локализованы в I зоне – основании ствола, а в продолговатом мозге они проходят в составе пирамид.
Экстрапирамидные пути начинаются от ядер серого вещества, расположенных в стволе головного мозга. Они проводят импульсы, обеспечивающие бессознательные (непроизвольные) движения и поддержание тонуса мышц. Эти тракты проходят во II зоне ствола головного мозга – покрышке.
Пирамидные пути
Основными эфферентными пирамидными путями являются корково-спинномозговой и корково-ядерный тракты.
1. Корково-спинномозговой путь проводит сознательные (волевые) двигательные нервные импульсы, обеспечивающие управление скелетной мускулатурой туловища и конечностей, выполнение точных высокодифференцированных движений. Кроме того, этот путь проводит тормозные импульсы от коры полушарий большого мозга к нейронам двигательных ядер передних рогов спинного мозга (рис. 4.9), т.е. он оказывает тормозное воздействие на сегментарный аппарат спинного мозга.
Корково-спинномозговой путь образован аксонами пирамидных клеток коры предцентральной извилины и околоцентральной дольки. Часть аксонов направляется из постцентральной извилины.

Рис. 4.9. Корково-спинномозговые пути:
1 – перекрест пирамид; 2 – латеральный корковоспинномозговой путь; 3 – латеральный канатик; 4 – передний корешок спинномозгового нерва; 5 – двигательные ядра передних рогов спинного мозга; 6 – передний канатик; 7 – передний рог; 8 – передний корково-спинномозговой путь; 9 – продолговатый мозг; 10 – корково-спинномозговой путь; 11 – внутренняя капсула; 12 – клетки Беца
В коре предцентральной извилины пирамидные нейроны локализуются по правилу "моторный гомункулюс Пенфилда". В самых верхних отделах предцентральной извилины располагаются нейроны, которыми начинаются эфферентные пути для иннервации мышц нижней конечности, причем в парацентральной дольке находится соматотопическая проекция мышц стопы, латеральнее – мышц голени и бедра. Далее располагаются нейроны, дающие начало эфферентным нервным путям к мускулатуре туловища. Среднюю треть предцентральной извилины занимают нейроны, обеспечивающие иннервацию мышц верхней конечности (вверху для мышц плеча, ниже – предплечья и кисти). Следует отметить, что площадь соматотопических проекционных зон в коре полушарий пропорциональна сложности движений, выполняемых определенной группой мышц. Наибольшую по площади соматотопическую проекцию имеют мышцы кисти (см. рис. 3.26).
Корково-спинномозговой тракт идет в нисходящем направлении во внутреннюю капсулу, в которой занимает переднюю часть задней ножки. Расположение волокон этого тракта во внутренней капсуле имеет характерные особенности: в непосредственной близости к колену внутренней капсулы проходят волокна, проводящие нервные импульсы для мышц верхней конечности, позади них – волокна для мышц туловища и, наконец, – волокна для мышц нижней конечности.
Далее корково-спинномозговой путь проходит по вентральной поверхности ствола головного мозга. В мосту он рассыпается на большое количество мелких пучков, отделенных друг от друга ядрами моста. В области продолговатого мозга разрозненные пучки волокон вновь собираются в один крупный пучок, который проходит в составе пирамиды. На границе между продолговатым и спинным мозгом большая часть волокон каждой пирамиды переходит на противоположную сторону (80%), образуя с аналогичными волокнами противоположной стороны перекрест пирамид. Из состава пирамиды 20% волокон остаются на своей стороне и продолжаются в передний канатик спинного мозга. Они составляют передний корково-спинномозговой путь. Перекрещенные волокна направляются в боковой канатик спинного мозга. В боковом канатике это наиболее крупный пучок волокон, он носит название латерального корково-спинномозгового пути. Таким образом, единый на протяжении ствола головного мозга корково-спинномозговой тракт в спинном мозге разделяется на два самостоятельных.
Латеральный корково-спинномозговой путь в каудальном направлении постепенно истончается. Наибольшее количество волокон от него отделяется в области утолщений спинного мозга, сегменты которых содержат эффекторные нейроны, отвечающие за иннервацию мышц верхних и нижних конечностей. Достигнув своего сегмента, волокна выходят из состава тракта и заканчиваются на клетках двигательных ядер передних рогов спинного мозга своей стороны.
Передний корково-спинномозговой путь располагается в переднем канатике спинного мозга. Он представляет собой сравнительно небольшой пучок волокон, основная часть которых посегментно в области передней белой спайки переходит на противоположную сторону и заканчивается на нейронах двигательных ядер передних рогов.
Аксоны вторых нейронов (мотонейронов), расположенных в двигательных ядрах передних рогов спинного мозга, покидают спинной мозг в составе передних корешков спинномозговых нервов. Затем они проходят в спинномозговых нервах и их ветвях к скелетной мускулатуре.
При поражении пирамидных нейронов и корково-спинномозговых путей возникают центральные параличи (выпадение двигательных функций) или парезы (ослабление двигательных функций). Центральный паралич характеризуется повышением тонуса парализованных мышц (гипертонус), повышением сухожильных рефлексов (гиперрефлексия), появлением патологических рефлексов. Эти проявления обусловлены отсутствием тормозного воздействия на сегментарный аппарат спинного мозга.
Если очаг поражения корково-спинномозгового пути локализуется на уровне верхних шейных сегментов, возникает паралич верхней и нижней конечностей на одноименной стороне. Если патологический очаг поражения находится в предцентральной извилине или в стволе головного мозга, возникает паралич конечностей на противоположной стороне, так как волокна корково-спинномозговых путей совершают перекрест.
При поражении периферического двигательного нейрона или его аксона возникает периферический паралич, который характеризуется атонией, арефлексией и атрофией. При этом движения полностью отсутствуют, а мышцы со временем атрофируются и замещаются жировой и соединительной тканями.
2. Корково-ядерный путь относится к группе двигательных пирамидных путей. Он проводит сознательные (волевые) двигательные нервные импульсы, обеспечивающие управление мускулатурой головы и частично шеи, выполнение точных и высокодифференцированных движений. Кроме того, этот путь проводит тормозные импульсы от коры полушарий большого мозга к нейронам двигательных ядер черепных нервов III, IV, V, VI, VII, IX, X, XI и XII пар, т.е. он оказывает тормозное воздействие на сегментарный аппарат ствола головного мозга.
Корково-ядерный путь образуется аксонами пирамидных клеток пятого слоя коры полушарий большого мозга. Большая часть аксонов происходит от клеток нижнелатеральной трети предцентральной извилины, меньшая часть – от клеток нижней трети постцентральной извилины. Участие в формировании корково-ядерного пути аксонов клеток нижней трети постцентральной извилины обусловлено соматотопической проекцией на кору полушарий жевательных и мимических мышц, мышц мягкого неба, глотки и гортани.
Аксоны пирамидных клеток веерообразно сходятся в пучок, который проходит через колено внутренней капсулы. Далее корково-ядерный путь проходит по вентральной поверхности ствола головного мозга – в средней части основания ножки мозга, основании моста и пирамидах продолговатого мозга.
В области среднего мозга от корково-ядерного пути отделяется часть волокон, которая заканчивается синапсами на клетках двигательных ядер глазодвигательного (III пара) и блокового (IV пара) черепных нервов, которые иннервируют мышцы глазного яблока.
В области моста от корково-ядерного пути вновь отделяются волокна, которые идут в дорсальном направлении и заканчиваются на нейронах двигательных ядер V, VI и VII пар черепных нервов. Аксоны мотонейронов двигательного ядра тройничного нерва иннервируют жевательные мышцы; челюстно-подъязычную и переднее брюшко двубрюшной мышцы; мышцу, напрягающую мягкое небо, а также мышцу, напрягающую барабанную перепонку. Аксоны клеток двигательного ядра отводящего нерва направляются к латеральной прямой мышце глазного яблока. Аксоны мотонейронов двигательного ядра VII пары иннервируют мимические мышцы, стременную мышцу, заднее брюшко двубрюшной мышцы, шилоподъязычную и подкожную мышцы.
Сравнительно небольшая часть волокон корково-ядерного пути достигает продолговатого мозга и верхних шейных сегментов спинного мозга. Эти волокна также отклоняются в дорсальном направлении и заканчиваются на нейронах двигательных ядер IX, X, XI и XII пар черепных нервов. Аксоны мотонейронов двойного ядра, общего для IX и X пар черепных нервов, иннервируют мышцы глотки, мягкого неба, гортани и верхней части пищевода. Аксоны мотонейронов двигательного ядра XI пары направляются к трапециевидной и грудино-ключично-сосцевидной мышцам, а аксоны мотонейронов двигательного ядра XII пары идут к мышцам языка.
Одностороннее разрушение пирамидных нейронов в нижнем отделе предцентральной извилины или поражение корково-ядерного пути вызывают не паралич, а парез (ограничение произвольных движений и снижение сократительной силы мышц), так как мотонейроны двигательных ядер черепных нервов в большинстве случаев получают нервные импульсы из обоих полушарий. Исключение составляют мышцы языка и мимические мышцы. К нейронам двигательного ядра подъязычного нерва идут только перекрещенные волокна корково-ядерного пути, поэтому их поражение вызывает паралич мышц языка с противоположной стороны. Мотонейроны двигательного ядра лицевого нерва, связанные с иннервацией нижней половины лица, получают только перекрещенные волокна.
Мотонейроны, связанные с иннервацией мускулатуры верхней половины лица, получают волокна от корково-ядерных путей своей и противоположной сторон. В связи с этим полный паралич мускулатуры развивается только в нижней половине лица на стороне, противоположной очагу поражения. В верхней половине лица отмечается лишь парез мимических мышц. Только двустороннее поражение корковых центров или корково-ядерных путей приводит к развитию центрального паралича.
При разрушении всех мотонейронов двигательных ядер черепных нервов или повреждении их аксонов возникает периферический паралич, который приводит к исчезновению рефлексов (арефлексия), утрате тонуса мышц (атония) и их атрофии.
Система эфферентных нейронов, тела которых располагаются в коре большого мозга, оканчиваются в двигательных ядрах черепных нервов и сером веществе спинного мозга. В составе пирамидного пути (tractus pyramidalis) выделяют корково-ядерные волокна (fibrae corticonucleares) и корково-спинномозговые волокна (fibrae corticospinales). И те, и другие являются аксонами нервных клеток внутреннего, пирамидного, слоя коры большого мозга (Кора большого мозга). Они располагаются в предцентральной извилине и прилегающих к ней полях лобной и теменной долей. В предцентральной извилине локализуется первичное двигательное поле, где располагаются пирамидные нейроны, управляющие отдельными мышцами и группами мышц. В этой извилине существует соматотопическое представительство мускулатуры. Нейроны, управляющие мышцами глотки, языка и головы, занимают нижнюю часть извилины; выше располагаются участки, связанные с мышцами верхней конечности и туловища; проекция мускулатуры нижней конечности находится в верхней части предцентральной извилины и переходит на медиальную поверхность полушария.
Пирамидный путь образуют преимущественно тонкие нервные волокна, которые проходят в белом веществе полушария и конвергируют к внутренней капсуле (рис. 1). Корково-ядерные волокна формируют колено, а корково-спинномозговые волокна — передние 2 /3 задней ножки внутренней капсулы. Отсюда пирамидный путь продолжается в основание ножки мозга и далее в переднюю часть моста (см. Головной мозг). На протяжении ствола мозга корково-ядерные волокна переходят на противоположную сторону к дорсолатеральным участкам ретикулярной формации, где они переключаются на двигательные ядра III, IV, V, VI, VII, IX, X, XI, XII черепных нервов (Черепные нервы); только к верхней трети ядра лицевого нерва идут неперекрещенные волокна. Часть волокон пирамидного пути проходит из ствола головного мозга в мозжечок.
В продолговатом мозге пирамидный путь располагается в пирамидах, которые на границе со спинным мозгом образуют перекрест (decussatio pyramidum). Выше перекреста пирамидный путь содержит от 700 000 до 1 300 000 нервных волокон с одной стороны. В результате перекреста 80% волокон переходит на противоположную сторону и образует в боковом канатике спинного мозга (Спинной мозг) латеральный корково-спинномозговой (пирамидный) путь. Неперекрещенные волокна из продолговатого мозга продолжаются в передний канатик спинного мозга в виде переднего корково-спинномозгового (пирамидного) пути. Волокна этого пути переходят на противоположную сторону на протяжении спинного мозга в его белой спайке (посегментно). Большинство корково-спинномозговых волокон оканчивается в промежуточном сером веществе спинного мозга на его вставочных нейронах, лишь часть их образует синапсы непосредственно с двигательными нейронами передних рогов, которые дают начало двигательным волокнам спинномозговых нервов (Нервы). В шейных сегментах спинного мозга оканчивается около 55% корково-спинномозговых волокон, в грудных сегментах 20% и в поясничных сегментах 25%. Передний корково-спинномозговой путь продолжается только до средних грудных сегментов. Благодаря перекресту волокон в П. с. левое полушарие головного мозга управляет движениями правой половины тела, а правое полушарие — движениями левой половины тела, однако мышцы туловища и верхней трети лица получают волокна пирамидного пути из обоих полушарий.
Функция П. с. состоит в восприятии программы произвольного движения и проведении импульсов этой программы до сегментарного аппарата ствола головного и спинного мозга.
Патология. Нарушения функции П. с. наблюдаются при многих патологических процессах. В нейронах П. с и их длинных аксонах нередко возникают нарушения обменных процессов, которые приводят к дегенеративно-дистрофическим изменениям этих структур. Нарушения бывают генетически обусловленными или являются следствием интоксикации (эндогенной, экзогенной), а также вирусного поражения генетического аппарата нейронов. Для дегенерации характерно постепенное, симметричное и нарастающее расстройство функции пирамидных нейронов, в первую очередь, имеющих наиболее длинные аксоны, т.е. заканчивающихся у периферических мотонейронов поясничного утолщения. Поэтому пирамидная недостаточность в таких случаях вначале выявляется в нижних конечностях. К этой группе заболеваний относится семейная спастическая параплегия Штрюмпелля (см. Параплегия), портокавальная энцефаломиелопатия, Фуникулярный миелоз, а также синдром Миллса — односторонний восходящий паралич неясной этиологии. Он начинается обычно в возрасте от 35—40 до 60 лет центральным парезом дистальных отделов нижней конечности, который постепенно распространяется на проксимальные отделы нижней, а затем и на всю верхнюю конечность и переходит в спастическую гемиплегию с вегетативными и трофическими нарушениями в парализованных конечностях. П. с. поражается часто при медленных вирусных инфекциях, таких как Амиотрофический боковой склероз, Рассеянный склероз и др. Почти всегда в клинической картине очаговых поражений головного и спинного мозга имеются признаки нарушения функции пирамидной системы. При сосудистых поражениях головного мозга (кровоизлияние, ишемия) пирамидные расстройства развиваются остро или подостро с прогрессированием при хронической недостаточности мозгового кровообращения. П. с. может вовлекаться в патологический процесс при энцефалитах (Энцефалиты) и Миелитах, при черепно-мозговой травме (Черепно-мозговая травма) и позвоночно-спинномозговой травме (Позвоночно-спинномозговая травма), при опухолях центральной нервной системы и др.
При поражении П. с. развиваются центральные парезы и Параличи с характерными нарушениями произвольных движений. Повышаются мышечный тонус по спастическому типу (трофика мышц обычно не изменяется) и глубокие рефлексы на конечностях, снижаются или исчезают кожные рефлексы (брюшные, кремастерные), появляются патологические рефлексы на руках — Россолимо — Вендеровича, Якобсона — Ласка, Бехтерева, Жуковского, Гоффманна, на ногах — Бабинского, Оппенгейма, Чаддока, Россолимо, Бехтерева и др. (см. Рефлексы). Характерным для пирамидной недостаточности является симптом Жюстера: укол булавкой кожи в области возвышения большого пальца руки вызывает сгибание большого пальца и приведение его к указательному при одновременном разгибании остальных пальцев и тыльном сгибании кисти и предплечья. Нередко выявляется симптом складного ножа: при пассивном разгибании спастичной верхней конечности и сгибании нижней конечности исследующий испытывает вначале резкое пружинящее сопротивление, которое затем внезапно ослабевает. При поражении П. с. часто отмечаются глобальные, координаторные и имитационные Синкинезии.
Для выявления пареза легкой степени используют специальные тесты: двигательный ульнарный рефлекс Вендеровича (возможность отвести мизинец от безымянного пальца при его щипкообразном оттягивании), проба Барре (при закрытых глазах поднять вверх и фиксировать обе руки ладонями внутрь с растопыренными пальцами; на стороне пирамидной недостаточности рука быстрее устает, опускается и сгибается в локтевом и лучезапястном суставах) и др.
В зависимости от локализации патологического очага и его размеров наблюдаются различные варианты клинической картины нарушения функции П. с. При поражении коры предцентральной извилины развивается спастическая моноплегия — паралич одной верхней или нижней конечности, кисти или стопы или даже отдельных пальцев. Возможно изолированное поражение пирамидных нейронов, связанных с ядрами лицевого и подъязычного нервов. При этом развивается центральный паралич (парез) мимической мускулатуры нижней части лица, имеющей одностороннюю надъядерную иннервацию, и мышц одной половины языка. При обширном корковом очаге на наружной поверхности предцентральной извилины развивается фациолингвобрахиальный паралич. При раздражении сохранных нейронов возникают пароксизмы джексоновской эпилепсии (Джексоновская эпилепсия) или кожевниковской эпилепсии (Кожевниковская эпилепсия).
Наиболее часто П. с. поражается на уровне внутренней капсулы. При этом развивается спастический паралич конечностей на стороне, противоположной патологическому очагу. Для такой капсулярной гемиплегии характерна полная утрата произвольных движений в дистальных отделах конечностей (в кисти и стопе), типичной является поза Вернике — Манна (рис. 2): рука согнута в локтевом, лучезапястном и межфаланговых суставах, приведена к туловищу, бедро и голень разогнуты, стопа в положении подошвенного сгибания и повернута внутрь. При ходьбе паретичная нога выносится вперед, описывая полукруг (так называемая косящая походка). Спастичность, а затем и контрактура парализованных мышц возникают вследствие одновременного поражения во внутренней капсуле аксонов пирамидной и экстрапирамидной систем.
Разрушение пирамидной системы на уровне ствола головного мозга проявляется различными альтернирующими синдромами (Альтернирующие синдромы): Вебера (при поражении одной ножки мозга), Фовилля или Мийяра — Гюблера (при поражении половины моста мозга), Авеллиса, Джексона (при поражении продолговатого мозга) и др.
При разрушении спинного мозга выше шейного утолщения развивается спастическая тетраплегия, а ниже шейного утолщения (но выше поясничного утолщения) — спастическая параплегия. Двустороннее поражение корково-ядерных трактов приводит к возникновению псевдобульбарного паралича (Псевдобульбарный паралич) (пареза), одной половины поперечника спинного мозга (правой или левой) — к развитию Броун-Секара синдрома (Броун — Секара синдром).
Диагноз поражения П. с. устанавливают на основании исследования движений больного и выявления признаков пирамидной недостаточности (наличие пареза или паралича, повышения мышечного тонуса, повышения глубоких рефлексов, клонусов, патологических кистевых и стопных знаков), особенностей клинического течения и результатов специальных исследований (электронейромиографии, электроэнцефалографии, томографии и др.).
Дифференциальный диагноз пирамидных параличей проводят с периферическими парезами и параличами, которые развиваются при поражении периферических мотонейронов. Для последних характерны гипотрофия и атрофия паретичных мышц, снижение мышечного тонуса (гипо- и атония), ослабление или отсутствие глубоких рефлексов, изменения электровозбудимости мышц и нервов (реакция перерождения). При остром развитии поражения П. с. в первые несколько часов или суток нередко наблюдается снижение мышечного тонуса и глубоких рефлексов в парализованных конечностях. Связано это с состоянием Диашиза, после устранения которого наступает повышение мышечного тонуса и глубоких рефлексов. Вместе с тем пирамидные знаки (симптом Бабинского и т.п.) выявляются и на фоне признаков диашиза.
Лечение поражений П. с. направлено на основное заболевание. Применяют лекарственные препараты, улучшающие метаболизм в нервных клетках (ноотропил, церебролизин, энцефабол, глутаминовую кислоту, аминалон), проведение нервного импульса (прозерин, дибазол), микроциркуляцию (вазоактивные препараты), нормализующие мышечный тонус (мидокалм, баклофен, лиоресал), витамины группы В, Е. Широко используют ЛФК, массаж (точечный) и рефлексотерапию, направленные на снижение мышечного тонуса; физио- и бальнеотерапию, ортопедические мероприятия. Нейрохирургическое лечение проводится при опухолях и травмах головного и спинного мозга, а также при ряде острых нарушений мозгового кровообращения (при тромбозе или стенозе экстрацеребральных артерий, внутримозговой гематоме, мальформациях церебральных сосудов и др.).
Библиогр.: Блинков С.М. и Глезер И.И. Мозг человека в цифрах и таблицах, с. 82, Л., 1964; Болезни нервной системы, под ред. П.В. Мельничука, т. 1, с. 39, М., 1982; Гранит Р. Основы регуляции движений, пер с англ., М., 1973; Гусев Е.И., Гречко В.Е. и Бурд Г.С. Нервные болезни, с. 66, М., 1988; Дзугаева С.Б. Проводящие пути головного мозга человека (в онтогенезе), с. 92, М., 1975; Костюк П.К. Структура и функция нисходящих систем спинного мозга, Л. 1973; Лунев Д.К. Нарушение мышечного тонуса при мозговом инсульте, М. 1974; Многотомное руководство по неврологии, под ред. Н.И. Гращенкова, т. 1, кн. 2, с. 182, М., 1960; Скоромец Д.Д. Топическая диагностика заболеваний нервной системы, с. 47, Л., 1989; Турыгин В.В. Проводящие пути головного и спинного мозга, Омск. 1977.
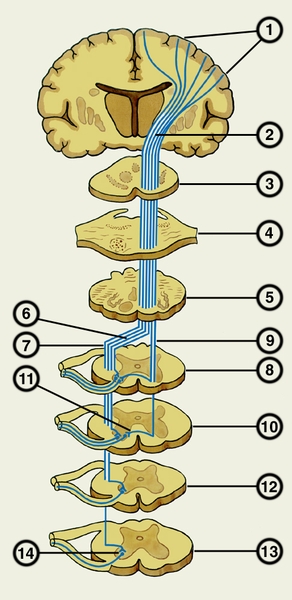
Рис. 1. Схематическое изображение пирамидного пути на различных уровнях головного и спинного мозга: 1 — пирамидные нейроны коры большого мозга; 2 — внутренняя капсула; 3 — средний мозг; 4 — мост; 5 — продолговатый мозг; 6 — перекрест пирамид; 7 — латеральный корково-спинномозговой (пирамидный) путь; 8, 10 — шейные сегменты спинного мозга; 9 — передний корково-спинномозговой (пирамидный) путь; 11 — белая спайка; 12 — грудной сегмент спинного мозга; 13 — поясничный сегмент спинного мозга; 14 — двигательные нейроны передних рогов спинного мозга.

Рис. 2. Поза Вернике — Манна и шаговое движение парализованной нижней конечности при левостороннем гемипарезе.
Читайте также:
