
Рецепторы мышечно суставной чувствительности
Проприорецепторы являются исполнительным отделом мышечно-суставной рецепции. Это механорецепторы, отслеживающие информацию о положении и смещении различных частей тела.
Благодаря проприорецепторам, осуществляется координация подвижных систем в состоянии покоя и во время двигательных актов. Мышечные веретена, сухожильные рецепторы Гольджи и суставные рецепторы составляют проприорецепторы человека.
Мышечные веретена –образования с афферентными и эфферентными нервными волокнами. В зависимости от сложности функционального назначения мышцы количество в ней мышечных веретен различно. В глубоких мышцах шеи их 63 на 1 г веса мышцы, в мышцах бедра – 5.
Каждое веретено состоит из тонких интрафузальных (внутриверетенных) поперечно-полосатых мышечных волокон и экстрафузальных (в 2 – 3 раза тоньше обычных волокон скелетных мышц). Длинные, толстые интрафузальные волокна отвечают за динамический компонент движения, короткие, тонкие – за статический.
Один конец мышечного веретена прикрепляется к сухожилию, другой – к мышечным волокнам. При растянутой мышце происходит растяжение мышечных веретен, и возникают нервные импульсы, идущие в ЦНС. При сокращении мышцы – натяжение мышечных веретен ослабевает, и поток импульсов заканчивается.
С увеличением степени и скорости растяжения мышцы проприорецептивная импульсация в ЦНС учащается. Импульсы от мышечных веретен продолжаются весь период нахождения мышцы в растянутом состоянии. Соответствующая импульсация информирует ЦНС о состоянии длины мышцы. В целом импульсация от рецепторов определяется длиной мышц, скоростью и силой её растяжения. При подготовке к движению и в его динамике происходит повышение чувствительности веретен.
Сухожильные рецепторы Гольджи –расположены в месте соединения мышечных волокон с сухожилием. Образуют сеть тонких сухожильных волокон, адекватно реагируют на напряжение мышцы и передают информацию в ЦНС о силе их напряжения, однако слабо реагируют на растяжение мышцы.
Суставные рецепторы –свободные или заключённые в специальную капсулу нервные окончания, отслеживают положение сустава в плане изменения суставного угла и скорости движения. Передают возникающие импульсы в кору больших полушарий.
Кинетические сигналы, поступающие от рецепторов мышечных веретен, сухожильных органов, суставных рецепторов, тактильных рецепторов кожи, информируют ЦНС о движении отдельной части тела. Эти же рецепторы составляют функциональное звено других органов чувств. В частности при оценке органом зрения расстояния до какого-либо предмета большая роль принадлежит проприорецептивной информации.
Проприорецептивные сигналы мышечной сенсорной системы передают импульсы биполярным нейронам спинномозговых узлов, один отросток которых связан с рецепторами, другой – заходит в спинной мозг и передаёт импульсы вторым нейронам в бульбарный отдел мозга (часть проводящих путей направляется в кору мозжечка), далее – к релейным ядрам таламуса промежуточного мозга и заканчивается в передней центральной извилине коры больших полушарий.
Зрительная рецепция
Зрительная сенсорная система включает глаз с оптической системой, фокусирующей световые лучи, обеспечивающей изображение на сетчатке.
Сетчаткасодержит фоторецепторы, тела биполярных и ганглиозных нейронов, зрительный нерв, несущий информацию нейронам переднего двухолмия среднего мозга и ядрам промежуточного мозга (наружные коленчатые тела), в проекционные поля затылочной области коры больших полушарий, где возникает ощущение, опознание и осмысление зрительного образа. Третичные поля обеспечивают взаимосвязь зрительной информации с другими сенсорными системами.
Глазное яблокосостоит из трёх оболочек, окружающих его внутреннее ядро: фиброзной (наружной), сосудистой (средней) и сетчатой (внутренней).
а) Фиброзная оболочка играет защитную роль. В заднем отделе (две трети оболочки) она образует белочную оболочку или склеру, в переднем (одна треть) –прозрачную роговицу.
Белочная оболочка содержит эластические и коллагеновые волокна, в нём мало основного вещества. Волокна образуют плотную пластинку, в наружном слое которой нет пигментных клеток. На медиальной части заднего полюса глаза она имеет решётчатое строение. Через рёшетчатые отверстия проникают отростки нервных клеток, формирующие зрительный нерв. На границе с роговицей в толще склеры проходит круговой венозный – шлемов канал.
Роговица –прозрачная, округлая, выпуклая кпереди и вогнута сзади пластинка (подобие часового стекла). Содержит много чувствительных нервных окончаний.
б) Сосудистая оболочка содержит сплетения кровеносных сосудов и пигментных клеток. Разделяется на три части: радужную оболочку, ресничное тело, собственно сосудистую оболочку.
Радужная оболочкаимеет вид круговой, вертикально стоящей пластинки с круглым отверстием – зрачком, выполняющим роль диафрагмы. При сильном свете зрачок суживается, при слабом – расширяется, что обеспечивается сокращением радиальных и циркуляторных мышечных волокон. Первые иннервируются симпатическими волокнами, вторые – парасимпатическими. Передняя поверхность оболочки имеет различную окраску, обуславливая цвет глаз.
Ресничное телов форме циркулярного валика располагается в области перехода склеры в роговицу. На внутренней поверхности валика имеются радиально ориентированные отростки. От них отходят ресничные связки, которые вплетаются в капсулу хрусталика. Процесс аккомодации (приспособления глаза к близкому или дальнему видению) возможен благодаря ослаблению или натяжению ресничных связок, находящихся под контролем круговых и меридиональных мышц ресничного тела. При сокращении круговых мышц ресничные отростки приближаются к центру ресничного кружка и ресничные связки ослабляются. За счёт внутренней упругости хрусталика расправляется и увеличивается его кривизна, уменьшается фокусное расстояние (при рассмотрении близких предметов хрусталик становится более выпуклым). Одновременно с сокращением круговых мышечных волокон происходит сокращение и меридиональных.
Дальняя точка ясного видения для здорового глаза молодого человека лежит в бесконечности, лучи идущие от неё, фокусируются на сетчатке. Ближняя точка ясного видения находится на расстоянии 10 см от глаза. В старческом возрасте часть мышечных волокон ресничного тела замещается соединительной тканью, эластичность и упругость хрусталика уменьшается. Это приводит к нарушению зрения (ближняя точка видения в 60 лет отодвигается до 75 см).
Собственно сосудистая оболочка состоит из эластичных волокон, кровеносных и лимфатических сосудов, пигментных клеток. Она рыхло сращена с внутренней поверхностью белочной оболочки и легко смещается при аккомодации.
в) Сетчатая оболочка, сетчатка – внутренняя сосудистая оболочка глаза. Состоит из 11 слоёв, которые можно объединит в два листка: наружный (пигментный) и внутренний. Пигментный – образован эпителиальными клетками, содержащими меланосомы, придающие им чёрный цвет. Пигмент чёрного цвета поглощает попадающий в глаз свет, предотвращая его отражение и рассеивание. Этим обеспечивается создание чёткого отображения воспринимаемого объекта. Во внутреннем листке содержаться два вида вторичночувствующих, различных по функциональному значению фоторецепторов (палочки и колбочки), и несколько видов нервных окончаний.
На заднем полюсе глаза находится овальное пятно – диск зрительного нерва (слепое пятно – не содержит фоторецепторов). Латеральнее на 4 мм на уровне диска зрительного нерва лежит пятно с центральной ямкой, окрашенное в красно-жёлто- коричневый цвет. В пятне концентрируется фокус световых лучей – место наилучшего восприятия цветовых раздражителей. В пятне находится наибольшее скопление цветочувствительных клеток – колбочек.
Внутренне ядро глаза состоит из прозрачных светопреломляющих сред: стекловидного тела, хрусталика и водянистой влаги, наполняющей переднюю и заднюю глазные камеры.
а) Стекловидное тело – прозрачная масса, лежащая позади хрусталика в полости перед сетчаткой. Образовано прозрачным коллоидным веществом, состоящим из тонких редких соединительнотканных волокон, белков и гиалуроновой кислоты.
б) Хрусталик имеет вид двояковыпуклой линзы. Выпуклые части спереди и сзади – полюса, периферическая часть – экватор. Он заключён в прозрачную капсулу, о которой берут начало ресничные связки, заканчивающиеся в ресничном теле. Волокна этих связок, идущие от передней и задней частей капсулы хрусталика, соединяются вместе у ресничного тела.
в) Водянистая влага передней и задней камер глаза расположена в соответствующих камерах. Передняя камера ограничена внутренней поверхностью роговицы и передней поверхностью радужной оболочки, задняя камера – спереди задней поверхностью радужной оболочки, сзади – хрусталиком, сбоку – ресничным телом. Задняя камера сообщается через зрачковое отверстие с передней камерой. Жидкость камер служит питательным веществом для хрусталика и роговицы, а также участвует в преломлении лучей.
Фоторецепция
Фоторецепторы глазаделятся на два типа: палочки и колбочки.В сетчатке каждого глаза 6 – 7 млн. колбочек и 110 - 125 млн. палочек. Палочки и колбочки распределены в сетчатке по разному: центральную часть занимают колбочки, по направлению к периферии число их уменьшается, а количество палочек возрастает. В результате они занимают всю периферию сетчатки.
Палочки обладают большей, чем колбочки, чувствительностью и являются рецепторами преимущественно сумеречного зрения. Они содержат зрительный пигмент родопсин и воспринимают чёрно-белое изображение. Колбочки обеспечивают дневное зрение. В них содержится родственный родопсину пигмент йодопсин, чувствительный к красному, зелёному и синему цветам. Колбочковое зрение может быть цветным при условии наличия в сетчатке нескольких типов колбочек с разными зрительными пигментами, осуществляющими поглощение световых лучей в различных частях спектра. При нарушении поглощения лучей в определённых частях спектра наступаетцветовая слепота (дальтонизм).Чаще не различаются красный и зёлёный цвета.
Кнутри от слоя фоторецепторных клеток расположен слой биполярных нейронов. Изнутри к ним примыкает слой ганглиозных нервных клеток, отростки которых составляют волокна зрительного нерва. Возбуждение фоторецептора при действии света передаётся в зрительный нерв через биполярный нейрон и ганглиозную нервную клетку, образующих в местах контактов синапсы. Фоторецепторы через биполярные нейроны с одной ганглиозной клеткой образуют её рецептивное поле. Поля ганглиозных клеток связаны между собой и перекрывают друг друга. Соответствующая конструкция связи обеспечивает возможность одной ганглиозной клетке принимать информацию от тысячи фоторецепторов, т.е. высокую надёжность функционирования зрительной сенсорной системы по анализу и синтезу окружающей ситуации.
Восприятие пространства
Острота зрения.По способности глаза различать наименьшее расстояние между двумя точками определяет остроту зрения. Нормальный глаз различает две точки под углом в 60 с. Максимальной остротой зрения обладает жёлтое пятно. На периферии способность к пространственному различению в 4 раза ниже. В связи с этим центральное зрение отличается более высокой остротой. Остроту зрения определяют с помощью специальных таблиц. Нормальная острота принимается за единицу.
Поле зрения.Часть пространства, видимая глазом при фиксации взгляда в одной точке, называется полем зрения.
Границы поля зренияопределяют прибором – периметром, в современных условиях электронным. Для бесцветных предметов границы поля составляют: книзу – 70 о , кверху – 60 о , внутрь – 60 о , кнаружи – 90 о . Поля зрения для различных цветов меньше и снижаются от красного к зелёному цветам.
Совпадение поля зрения обоих глаз у человека весьма важно для восприятия пространства.
Зрение обеими глазами.При взгляде на предмет ощущения двух предметов не возникает. Это связано с попаданием изображения на общие участки двух сетчаток. В восприятии человека они сливаются в одно. Рассматривание близкого предмета, при конвергенции (схождении) глаз, изображение какой – либо отдалённой точки попадает на неидентичные (диспарантные) точки двух сетчаток. Диспарация играет большую роль в оценке расстояния и в видении глубины рельефа. Бинокулярное объединение сигналов от двух сетчаток в единый образ происходит в первичной зрительной коре.
Оценка расстояния.Оценка глубины пространства, т.е. расстояния до рассматриваемого предмета возможна при зрении одним глазом (монокулярное зрение) и двумя глазами (бинокулярное зрение), которое точнее. В оценке близких расстояний при монокулярном зрении имеет место явление аккомодации (изменение формы хрусталика).
Оценка величины объекта.В оценке величины предмета используются величина изображения на сетчатке и расстояние предмета от глаза. Грубые ошибки в определении величины предмета связаны с трудностью оценки расстояния, на котором он находится.
Вопросы к семинару
2. Классификация рецепторов: а) по характеру взаимодействия раздражителей; б) в зависимости от физической природы раздражителей.
3.Классификация рецепторов: в) в зависимости от контакта с раздражителем; г) по структурным особенностям и принципу преобразования энергии раздражения.
4.Чувствительность сенсорной системы.
5.Адаптационные процессы сенсорных систем.
6. Кожная рецепция.
7. Болевая и температурная чувствительность.
8. Тактильная чувствительность.
9. Висцерорецептивная сенсорная система.
10. Мышечно-суставная чувствительность (проприорецепция).
11. Зрительная рецепция. Сетчатка.
12. Зрительная рецепция. Глазное яблоко (фиброзная оболочка).
13. Зрительная рецепция. Глазное яблоко (сосудистая оболочка).
14. Зрительная рецепция. Глазное яблоко (сетчатая оболочка).
15. Зрительная рецепция. Внутреннее ядро глаза.
Пути, по которым чувствительные импульсы достигают коры большого мозга, состоят из трех нейронов. Клетки первого нейрона всех видов чувствительности лежат в спинномозговых узлах (ганглиях) или в их аналогах—чувствительных ядрах черепных нервов (V, VIII, IX, X пары). Периферический отросток чувствительной клетки в составе периферического нерва идет в кожу, к мышцам, связкам, сухожилиям; центральный отросток через задний корешок вступает в спинной мозг.
В спинном мозге различные виды чувствительности проводятся вверх по-разному. Волокна болевой, температурной и частично тактильной чувствительности вступают в задние рога спинного мозга. Здесь происходит переключение импульса на второй нейрон.
Аксоны второго нейрона совершают на своем уровне перекрест и входят в боковой канатик спинного мозга противоположной стороны, образуя латеральный спиноталамический путь, который поднимается вверх, проходит через ствол мозга и заканчивается в латеральном вентральном ядре таламуса, где расположены клетки третьего чувствительного нейрона.

Волокна глубокой и частично тактильной чувствительности, войдя в спинной мозг, минуют задние рога и направляются непосредственно в задние столбы —в пучки Голля (тонкий пучок) и Бурдаха (клиновидный пучок). Тонкий пучок несет импульсы от нижней половины туловища и ног, клиновидный — от верхней половины туловища и рук.
Эти пучки поднимаются вверх до продолговатого мозга, в дорсальных отделах которого лежат клетки второго нейрона глубокой чувствительности. Аксоны вторых нейронов на уровне олив моста мозга совершают перекрест, образуя медиальную петлю (lemniscus medialis), присоединяются к пути поверхностной чувствительности и вступают в латеральное вентральное ядро зрительного бугра (третий нейрон).
От этого ядра волокна всех видов чувствительности идут через задний отдел задней ножки внутренней капсулы и далее восходят в теменную долю мозга, главным образом в область постцентральной извилины (поля 1, 2, 3, 5, 7). Здесь все чувствительные пути оканчиваются у корковых клеток, которые расположены преимущественно во втором и четвертом слоях коры (четвертый нейрон).
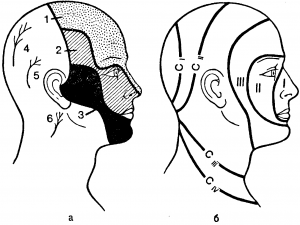
Чувствительная иннервация лица, отчасти оболочек и сосудов мозга осуществляется тройничным нервом, первые нейрон которого находится в тройничном (гассеровом) узле. Последний лежит в углублении на передней поверхности височной кости. В клетках тройничного узла есть сомато-топика, т. е. клетки поверхностных слоев связаны с задними отделами лица, глубоких —с передними.
Периферические отростки клеток этого узла образуют три ветви. Первая ветвь, выходя из полости черепа через верхнюю глазничную щель (fissura orbitalis superior), иннервирует область лба и передней половины головы (рис. 1). Вторая ветвь покидает череп через круглое отверстие (foramen rotandum) и иннервирует среднюю часть лица и верхнюю челюсть.
Третья ветвь выходит из полости черепа через овальное отверстие (foramen ovale) и осуществляет иннервацию лица на уровне нижней челюсти. Центральные отростки клеток тройничного узла, войдя в мост мозга, делятся на два пучка. Волокна болевой, температурной и частично тактильной чувствительности оканчиваются в ядре спинномозгового пути тройничного нерва (nucl. spinalis) — аналоге заднего рога спинного мозга, волокна мышечно-суставного и частично тактильного чувства — в мостовом ядре тройничного нерва (nucl. pontinus) (аналог второго нейрона глубокой чувствительности).
Аксоны вторых нейронов поднимаются вверх до зрительного бугра, где после переключения на третий нейрон восходят к коре в составе других чувствительных путей.
Иннервация глотки, гортани, надгортанника, барабанной полости и наружного слухового прохода осуществляется чувствительной порцией языкоглоточного и блуждающего нервов. Клетки первого чувствительного нейрона языкоглоточного и блуждающего нервов расположены в двух узлах: верхнем (gangl. superius) и нижнем (gangl. inferius).
Периферические отростки клеток этих узлов иннервируют глотку, слуховую (евстахиеву) трубу, барабанную полость, наружный слуховой проход. Центральные отростки вступают в продолговатый мозг и заканчиваются в общем для обоих нервов чувствительном ядре —nucl. alae cinereae. В этом ядре расположены клетки второго нейрона.
Его аксоны поднимаются к зрительному бугру, где лежат клетки третьего нейрона. Отсюда в составе всех чувствительных путей аксоны третьего нейрона идут в кору большого мозга— к клеткам нижнего отдела задней центральной извилины (четвертый нейрон). Наиболее важны следующие анатомо-физиологические особенности общей чувствительности.
Специфика различных видов чувствительности (тепло, холод, боль) обусловлена различным строением рецепторного аппарата на периферии. Строение периферических нервных окончаний в коже, мышцах, сухожилиях чрезвычайно разнообразно (свободные окончания, тельца Мейсснера, тельца Краузе, тельца Руффини).
Свободные окончания воспринимают болевые раздражения, тельца Краузе — холодовые, тельца Руффини — тепловые, тельца Мейсснера — тактильные. Вместе с тем специфичность этих рецепторных аппаратов, по-видимому, не абсолютная, так как ощущение боли можно получить раздражением не только свободных окончаний, но и телец Мейсснера, Руффини и др.
По миелинизированным волокнам в кору поступают в основном импульсы глубокой и тактильной чувствительности, по тонкомиелиновым и безмиелиновым — болевой и температурной. Болевые ощущения проводятся и миелинизированными волокнами — они дают быстро нарастающее локальное чувство боли. Раздражение безмиелиновых волокон вызывает менее четко локализованную боль, нередко с неприятным компонентом.
Несмотря на тесное анатомическое единство всех чувствительных систем, сохраняется раздельное проведение болевого, температурного, тактильного и мышечно-суставного ощущений вплоть до подкорки и коры большого мозга.
В заднем корешке миелиновые волокна лежат медиально, тонкомиелиновые и безмиелиновые — более латерально; во внутренней капсуле медиально располагаются проводники, несущие глубокую чувствительность, более латерально — проводники, несущие тактильную, температурную, болевую чувствительность. Вследствие этого возможно более или менее изолированное выпадение поверхностной или глубокой чувствительности при очагах на различных уровнях.
Лоскутность зон периферической чувствительной иннервации обусловлена тем, что афферентные импульсы идут в составе смешанных периферических нервов, осуществляющих также двигательную и вегетативную функции.
В области заднего корешка и заднего рога происходит топическое упорядочение, систематизация чувствительных импульсов, поступающих с периферии, т. е. информация принимается группами клеток со строго очерченных сегментарных зонпродольных на конечностях (в виде лампаса) и поперечных на туловище (в виде пояса).
Виды расстройств кожно мышечной чувствительности
Виды расстройств чувствительности многообразны. Самым частым симптомом поражения афферентных систем являются боли. Они различны по локализации и характеру, но наиболее выражены при поражении периферических нервов, задних корешков и зрительного бугра. В проведении болевых импульсов участвуют практически все афферентно-медиаторные системы (холинергическая, адренергическая, серотонинергическая, гистаминергическая и др.).
Разнообразные болевые ощущения являются универсальным предупредительным сигналом о повреждении или заболевании. Вместе с тем длительные или сильные боли необходимо купировать, хотя в самом организме (особенно в зрительном бугре и коре теменной доли большого мозга) в этих случаях происходит стимуляция образования веществ, блокирующих передачу или подавляющих боль, прежде всего энкефалинов и эндорфинов, взаимодействующих с опиатными рецепторами клеток.
Помимо медикаментозных средств (анальгетики, транквилизаторы, нейролептики, психостимуляторы), физиотерапии, электростимуляции, новокаиновых блокад, при резких, неподдающихся консервативному лечению болях иногда приходится прибегать к хирургическим вмешательствам.
Применяют операции: таламотамию (деструкция латерального вентрального ядра), трактотомию (перерезка чувствительных путей в продолговатом мозге), хордотомию (перерезка спиноталамического тракта; чаще ее производят на уровне верхнегрудных позвонков) и комиссуротомию (перерезка передней комиссуры; чаще оперируют на уровне нижнегрудных позвонков).
Эти оперативные вмешательства (их производят с использованием микрохирургической техники и методов стереотаксиса) приводят к стиханию или уменьшению болей.
Ощущение покалывания, ползания мурашек, онемения называется парестезией. Дизестезия — это извращенное восприятие раздражений, когда тактильное воспринимается как болевое, тепловое —как холодовое и т. д. Аллохейрия — патологическое восприятие раздражения, когда оно ощущается не в месте его нанесения, а в симметричной половине тела.
Полиестезия это вид извращения болевой чувствительности, при котором одиночное раздражение воспринимается как множественное. Анестезия — полная потеря чувствительности, гемианестезия — одной половины тела в области одной конечности — моноанестезией, в области ног и нижней части туловища — параанестезией. Гипестезия — понижение восприятия как всей чувствительности, так и отдельных ее видов.
Область выпадения может быть различной (гемигипестезия, моногипестезия). Гиперестезия — повышение чувствительности к различным видам раздражения вследствие понижения порога возбудимости. Гиперпатия — своеобразный вид чувствительных нарушений, который характеризуется тем, что любое, самое небольшое раздражение, если оно превышает порог возбудимости, сопровождается крайне неприятным ощущением боли и длительным последействием.
Сенестопатии — разнообразные тягостные, длительно беспокоящие больных ощущения жжения, давления, щекотания, стягивания и т.|п., не имеющих явных органических причин для их возникновения.
Боль, анестезия, парестезия и некоторые другие виды нарушений чувствительности могут появляться при вовлечении в процесс афферентных систем на различных уровнях (ганглии, задние корешки, спинной мозг, таламус и др.). Нарушение сложных видов чувствительности (чувство локализации, дискриминационная и кинестетическая чувствительность, двухмерно-пространственное чувство, стереогноз) обусловлены поражением высших корковых центров.
Статья на тему Кожно мышечная чувствительность
мышечно-суставного чувства (двигательный, или проприоцептивной анализатор). Этот анализатор имеет решающее значение в определении положения тела и его частей в пространстве, а также в обеспечении тонкой координации ции движений. Рецепторы мышечно-суставного чувства содержатся в мышцах, сухожилиях и суставах, называются проприорецепторов и к их числу относятся: тельца. Фатера-Пачини, голые нервные окончания, тельца. Г ольджи и мышечные веретена. По механизму действия все проприорецепторы относятся к механорецепторов. Тельца. Фатера-Пачини содержатся в сухожилиях, суставных сумках, фасциях мышц и периосте. Тельца. Гольд жи (цибулиноподибни тельца) представляют собой заполненную лимфой капсулу, в которую заходят сухожильные волокна, окруженные обнаженными нервными волоконцами (рис 19). Тельца. Гольджи (впервые описаны в 1880 г италийсь кем гистологом. К. Гольджи) обычно располагаются в сухожилияухожилках

(на границе мышечной и сухожильной ткани), а также в опорных участках капсул суставов и в суставных связках. Из рисунка ясно, что это рецепторное образования расположено"последовательно"в цепи"мышца - сухожилия"и, таким образом, его раздражение возникает при растяжении в этой цепи (например, при сокращении мышцы). Мышечные веретена это разделенными волокна длиной 1-4мм, окружены капсулой, которая по повнена лимфой (рис 20). В капсуле содержится от 3 до 13 так называемых интрафузальных волоконец. Количество мышечных веретен и содержание в них интрафузальных мышечных волокон в разных мышцах неодинаковы; чем ск ладниша работа выполняется мышцей, тем больше в нем рецепторных образований. Мышечные веретена отвечают. И на растяжение и на сокращение мышц, так как имеют двойную иннервацию: эфферентной и афференты нрентну.
Наличие двух рецепторных образований (телец. Гольджи и мышечных веретен) дает возможность получать тонко дифференцированную информацию о состоянии мышцы, т.е. степень его сокращения, расслабления или рас тягнення. Когда мышца расслаблена, происходит жидкая тоническая афферентная импульсация от сухожильных рецепторов. Гольджи и усиленная от мышечных веретен. При сокращении устанавливается противоположное спиввиднош ения, а при искусственном растяжениині


мышц афферентация усиливается от обоих видов рецепторов. Таким образом, любое состояние мышцы имеет свое отражение в характере импульсации от обоих видов рецепторов сухожильно-мышечных структур. Импульсы и, возникающих в проприорецепторов при движении, по центростремительным нервам поступают (через проводящие пути спинного и головного мозга) к мозжечка, ретикулярной формации, гипоталамуса и других стр уктур ствола мозга и далее-к сомато-сенсорных зон коры больших полушарий головного мозга, где и возникают ощущения изменения в положении частей тела в ответ на раздражение проприорецепторов привычно вине кають рефлекторные сокращения (расслабление) соответствующих групп мышц, или изменение их тонуса. Это способствует сохранению или изменению определенных движений, а также обусловливает поддержание позы и равновесия тела. При подъеме п редметив с помощью мышечно-суставного чувства можно приблизительно определить их весов вагу.
Кроме рассмотренного специфического сенсорного пути, импульсы от проприорецепторов влияют на деятельность многих внутренних органов, так как любая двигательная активность требует интенсификации поставки я кислорода, питательных веществ и удаления продуктов обмена веществ. Это в свою очередь требует усиления деятельности соответствующих внутренних органов в системах кровообращения, дыхания, выделения и др.. Такая согласованных енисть будет возможна, когда в вегетативных центров, регулирующих работу внутренних органов, будет поступать информация о состоянии мышцеів.
О сугубо сенсорную деятельность мышечного анализатора принято судить по точности восстановления положений суставов и ощущением изменения положения тела. Установлено, что наиболее чувствительным в этом смысле является пле ечовий сустав. Для него порог восприятия смещения при скорости 0,3 ° в секунду составляет 0,22-0,42 °. Наименее чувствителен голеностопный сустав, у которого порог составляет 1,15-1,30 °. В нормальном состоянии человек с закрытыми глазами обычно восстанавливает положение своего тела (с ошибкой до 3%) через 10-15 это5 сек.
У школьников возбудимость проприорецепторов с возрастом увеличивается: низкая она у учащихся 1 класса, самая высокая в учащихся XI класса. Главным условием нормального физического развития двигательных качеств детей является постоянная п поддержка активного состояния их проприорецепторов. Наибольшая нагрузка проприорецепторы получают в дни и часы уроков труда, физкультуры, занятий в спортивных секциях, игр и прогулок на улице, на йменше-в часы относительной недвижимости (на уроках, во время выполнения домашнего задания и пассивного отдыха). Активность рецепторов мышц повышается в первой половине дня и снижается две ечичір.
Проприоцепция — это группа сигналов, посылаемых в центральную нервную систему специальными терминалами (проприоцепторами), расположенными в суставных капсулах, связках, сухожилиях и мышцах.
Проприоцепторы, среди которых выделяют мышечные рецептор ных образований" href="http://www.braintools.ru/article/9580"> рецептор ы, или мышечные веретена ( рецептор ы растяжения), сухожильные рецептор ы, или органы Гольджи ( рецептор ы мышечной силы), а также суставные рецепторы относятся к механо рецептор ам, посылающим в ЦНС информацию о положении, деформации и смещениях различных частей тела.
Функционирование этих рецептор ов обеспечивает координацию всех подвижных органов и тканей животного и человека в состоянии покоя и во время любых двигательных актов. При экспериментальном выключении проприоцепторов животные теряют способность поддерживать естественные позы, двигаться и целесообразно реагировать на внешние воздействия.
Если человек закроет глаза и попытается написать текст, то буквы будут написаны достаточно четко. Этим простым способом легко убедиться в умении человека пользоваться информацией, идущей от мышц и суставов.
Проприоцепторы составляют периферическую часть проприоцептивной сенсорной системы , или двигательного анализатора . Вместе с тем, несмотря на то, что миллионы людей ежедневно пользуются услугами этого анализатора, мы до сегодняшнего дня знаем сравнительно мало о его деятельности. Это особенно касается работы коркового отдела двигательного анализатора. Внутренние проприоцепторы находятся в мышцах, сухожилиях, сухожильных влагалищах, межкостных мембранах, фасциях, тканях суставов, надкостнице и т.д. Среди них имеются неспециализированные рецептор ы, встречающиеся и в других частях тела (свободные нервные окончания, инкапсулированные рецептор ы типа телец Руффини и Пачини), и специализированные — мышечные веретена и сухожильные органы (или рецептор ы) Гольджи.
Двигательный, или кинестетический, анализатор (мышечная сенсорная система ) обеспечивает формирование так называемого мышечного чувства при изменении напряжения мышц, суставных сумок, связок и сухожилий. Проблема мышечно-суставных ощущений имеет исключительное значение для физиологии и психологии. Специфические особенности человека появились благодаря более совершенной организации мышечной сенсорной системы по сравнению с животными.
И.М. Сеченов считал, что мышечное чувство является ближайшим регулятором движений и одним из орудий ориентации человека в пространстве и времени. Ощущения положения и перемещения тела в пространстве, ощущения во время трудовой деятельности и членораздельная речь лежат в основе формирования сознания человека и его представлений об окружающем мире.
Мышечное чувство обладает тремя качествами. Это, во-первых, ощущение положения конечностей, когда человек может определить положение своих конечностей и их частей относительно друг друга. Во-вторых, ощущение движения, когда, изменяя угол сгибания в суставе, человек осознаёт скорость и направление движения. Третьим качеством является ощущение усилия, когда человек может оценить мышечную силу, нужную для движения или удерживания суставов в определённом положении при подъёме или перемещении груза.
Наряду с кожной, зрительной и вестибулярной сенсорными системами двигательный анализатор оценивает положение тела в пространстве, позу, участвует в координации мышечной деятельности.
Двигательная сенсорная система служит для анализа состояния двигательного аппарата — его движения и положения. Информация о степени сокращения скелетных мышц, натяжении сухожилий, изменении суставных углов необходима для регуляции двигательных актов и поз.
Общий план организации. Двигательная сенсорная система состоит из следующих 3-х отделов:
- периферический отдел, представленный проприо рецептор ами, расположенными в мышцах, сухожилиях и суставных сумках;
- проводниковый и отдел, который начинается биполярными клетками (первыми нейрон ами), тела которых расположены вне ЦНС — в спинномозговых узлах. Один их отросток связан с рецептор ами, другой входит в спинной мозги передает проприоцептивные импульсы ко вторым нейрон ам в продолговатый мозг (часть путей от проприо рецептор ов направляется в кору мозжечка ), а далее к третьим нейрон ам — релейным ядрам таламуса (в промежуточный мозг);
- корковый отдел находится в передней центральной извилине коры больших полушарий.
Функции проприо рецептор ов
К проприо рецептор ам относятся мышечные веретена, сухожильные органы (или органы Гольджи) и суставные рецептор ы ( рецептор ы суставной капсулы и суставных связок). Все эти рецептор ы представляют собой механо рецептор ы, специфическим раздражителем которых является их растяжение.
Мышечные веретена человека, представляют собой продолговатые образования длиной несколько миллиметров, шириной десятые доли миллиметра, которые расположены в толще мышцы. В разных скелетных мышцах число веретен на1 г ткани варьирует от нескольких единиц до сотни.
Таким образом, мышечные веретена как датчики состояния силы мышцы и скорости ее растяжения реагируют на два воздействия: периферическое — изменение длины мышцы, и центральное — изменение уровня активации гамма-мото нейрон ов. Поэтому реакции веретен в условиях естественной деятельности мышц довольно сложны. При растяжении пассивной мышцы наблюдается активация рецептор ов веретен; она вызывает миотатичес-кий рефлекс , или теор ии рефлекторной деятельности" href="http://www.braintools.ru/article/8998">рефлекс на растяжение. При активном сокращении мышцы уменьшение ее длины оказывает на рецептор ы веретена дезактивирующее действие, а возбуждение гамма-мото нейрон ов, сопутствующее возбуждению альфа-мото нейрон ов, приводит к реактивации рецептор ов. Вследствие этого импульсация от рецептор ов веретен во время движения зависит от длины мышцы, скорости ее укорочения и силы сокращения.
Сухожильные органы ( рецептор ы) Гольджи человека располагаются в зоне соединения мышечных волокон с сухожилием, последовательно по отношению к мышечным волокнам.
Сухожильные органы представляют собой структуру вытянутой веретенообразной или цилиндрической формы, длина которой у человека может достигать1 мм. Этот первичночувствующий рецептор . В условиях покоя, т.е. когда мышца не сокращена, от сухожильного органа идет фоновая импульсация. В условиях мышечного сокращения частота импульсации возрастает прямо пропорционально величине мышечного сокращения, что позволяет рассматривать сухожильный орган как источник информации о силе, развиваемой мышцей. В тоже время сухожильный орган слабо реагирует на растяжение мышцы.
В результате последовательного крепления сухожильных органов к мышечным волокнам (а в ряде случаев — к мышечным веретенам), растяжение сухожильных механо рецептор ов происходит при напряжении мышц. Таким образом, в отличие от мышечных веретен, сухожильные рецептор ы информируют нервные центры о степени напряжения мыши, и скорости его развития.
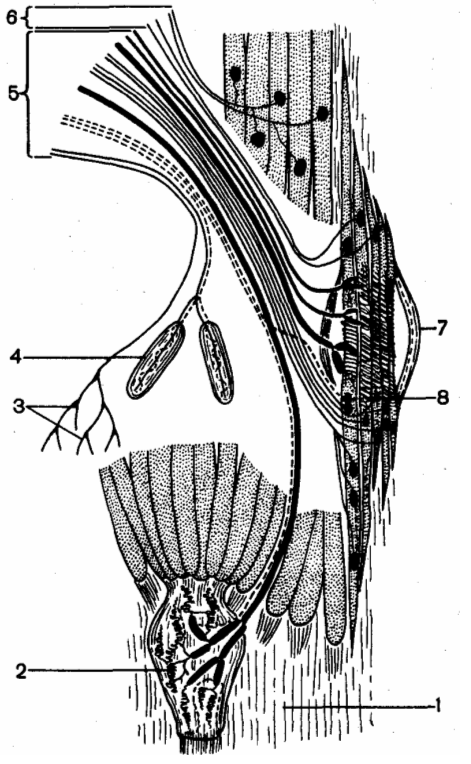
Эфферентная иннервация скелетной мышцы: 1 — сухожилие мышцы; 2 — сухожильные рецептор ы Гольджи; 3 — свободные окончания; 4 — тельца Фатера-Пачини; 5 — афферентные волокна; 6 — эфферентные волокна; 7 — мышечное веретено; 8 — окончания афферентных волокон
Суставные рецептор ы реагируют на положение сустава и на изменения суставного угла, участвуя, таким образом, в системе обратных связей от двигательного аппарата и в управлении им. Суставные рецептор ы информируют о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецептор ы представляют собой свободные нервные окончания или окончания, заключенные в специальную капсулу. Одни суставные рецептор ы посылают информацию о величине суставного угла, т. е. о положении сустава. Их импульсация продолжается в течение всего периода сохранения данного угла. Она тем большей частоты, чем больше сдвиг угла. Другие суставные рецептор ы возбуждаются только в момент движения в суставе, т. е. посылают информацию о скорости движения. Частота их импульсации возрастаете увеличением скорости изменения суставного угла.
Проводниковый и корковый отделы проприоцеп-тивного анализатора млекопитающих и человека. Информация от мышечных, сухожильных и суставных рецептор ов поступает через аксон ы первых афферентных нейрон ов, находящихся в спинномозговых ганглиях, в спинной мозг , где частично переключается на альфа-мото нейрон ы или на вставочные нейрон ы (например, на клетки Реншоу), а частично направляется по восходящим путям в высшие отделы головного мозга. В частности, по путям Флексига и Говерса проприоцептивная импульсация доставляется к мозжечку, а по пучкам Голля и Бурдаха, проходящим в дорсальных канатиках спинного мозга , она доходит до нейрон ов одноименных ядер, расположенных в продолговатом мозге.
Аксоны таламических нейрон ов ( нейрон ов третьего порядка) оканчиваются в коре больших полушарий, главным образом, в соматосенсорной коре (постцентральная извилина) и в области сильвие-вой борозды (соответственно, участки S-1 и S-2), а также частично в двигательной (префронтальной) области коры. Эта информация используется двигательными системами мозга достаточно широко, в том числе для принятия решения о замысле движения, а также для его реализации. Кроме того, у человека на основе про-приоцептивной информации формируются представления о состоянии мышц и суставов, а также, в целом, о положении тела в пространстве.
Сигналы, идущие от рецептор ов мышечных веретен, сухожильных органов, суставных сумок и тактильных рецептор ов кожи. Механо рецептор ная и температурная чувствительность" href="http://www.braintools.ru/article/9885"> рецептор ов кожи, называют кинестетическими, т. е. информирующими о движении тела. Их участие в произвольной регуляции движений различно. Сигналы от суставных рецептор ов вызывают заметную реакцию в коре больших полушарий и хорошо осознаются. Благодаря им человек лучше воспринимает различия при движениях в суставах, чем различия в степени напряжения мышц при статических положениях или поддержании веса. Сигналы же от других проприо рецептор ов, поступающие преимущественно в мозжечок, обеспечивают бессознательную регуляцию, подсознательный контроль движений и поз.
Таким, образом, проприоцептивные ощущения дают человеку возможность воспринимать изменения положения отдельных частей тела в покое и во время совершаемых движений. Информация, поступающая от проприоцепторов, позволяет ему постоянно контролировать позу и точность произвольных движений, дозировать силу мышечных сокращений при противодействии внешнему сопротивлению, например при подъеме или перемещении груза.
Проприоцептивное восприятие положения тела и движений происходит в результате объединения в соматосенсорной коре информации от всех разновидностей проприоцепторов.
Читайте также:
