
При растяжении мышечных волокон возбуждаются рецепторы
Следовательно, при управлении движениями высшие моторные уровни помимо прямых влияний на спинальные альфа-мотонейроны могут оказывать на них косвенное влияние - через гамма-петлю. Управление через гамма-петлю является важным механизмом сохранения положения звеньев тела и сохранения требуемых суставных углов. Дело в том, что при управлении, в котором используется спинальная гамма-петля, высшие моторные уровни освобождаются от непосредственного контроля за сохранением длины мышцы и, следовательно, за сохранением положения в каждом отдельном суставе. Им необходимо лишь обеспечить некоторый уровень гамма-активности и тем самым определить требуемую (эталонную) длину мышечных веретён (степень их сокращения). После этого сохранение требуемой длины мышцы может поддерживаться автоматически спинальным моторным уровнем с помощью механизма рефлекса на растяжение.
При осуществлении сложных двигательных актов происходит одновременная активация a- и g-мотонейронов: активируются a-мотонейроны (обеспечивая само движение) и g-мотонейроны (поддерживая возбуждение a-мотонейронов).
Рисунок 6. Рефлекторная регуляция напряжения мышцы
Таким образом, сухожильные рефлексы играют двоякую роль: 1. Участвуют в регуляции напряжения в мышце. Когда напряжение достигает значения, при котором возбуждаются сухожильные рецепторы Гольджи, a-мотонейроны, иннервирующие сокращающуюся мышцу, тормозятся, что приводит к ее расслаблению и падению в ней напряжения; 2. Способствуя быстрому расслаблению мышцы, они защищают её от повреждения при сильных сокращениях. Несмотря на это, при чрезмерных нагрузках и напряжении икроножной мышцы в ряде случаев у спортсменов и артистов балета может происходить отрыв ахиллова сухожилия от места прикрепления.
Запомните: В регуляции деятельности каждой мышцы участвуют две регулирующие системы обратной связи: система регуляции длины, роль датчика в которой играют мышечные веретёна, и система регуляции напряжения, датчиками которой служат сухожильные рецепторы.
Длина и напряжение мышцы взаимообусловлены. Если, например, вытянутая вперёд рука ослабит напряжение мышц, то уменьшится раздражение рецепторов Гольджи, а сила тяжести станет опускать руку. Это приведёт к растяжению мышц, нарастанию возбуждения интрафузальных рецепторов и соответствующей активации мотонейронов. В итоге произойдет сокращение мышц и рука вернётся в прежнее положение.
Сухожильные рефлексы названы так потому, что их можно вызвать, легко ударив неврологическим молоточком по сухожилию более или менее растянутой мышцы. От удара такая мышца растягивается и тотчас рефлекторно сокращается. В неврологической практике сухожильные рефлексы используются для оценки состояния соответствующих сегментов спинного мозга. Наиболее известный рефлекс растяжения - коленный, возникает в ответ на кратковременное растяжение четырехглавой мышцы бедра легким ударом по ее сухожилию ниже коленной чашечки. Расслабленная мышца растягивается, а возникшее в связи с этим возбуждение рецепторов мышечных веретён распространяется по рефлекторной дуге к этой же мышце. После короткого латентного периода происходит сокращение мышцы и разгибание конечности (рис. 7).
Рисунок 7. Схема рефлекторной
дуги коленного сухожильного рефлекса
Этот рефлекс отражает состояние III - IV поясничных сегментов спинного мозга, так как именно там расположены центры этого рефлекса. Кроме коленного рефлекса в клинике применяют еще целый ряд подобного рода рефлекторных проб. Например, постукивание в области локтевого сустава вызывает рефлекторное разгибание руки. Растяжению в этом случае подвергается трёхглавая мышца плеча.
Таким образом, самый низкий уровень в организации движения связан с двигательными системами спинного мозга. В спинном мозге имеются альфа-мотонейроны, которые прямо управляют мышцами; располагаются гамма-мотонейроны и вставочные нейроны (интеронейроны), образующие множество контактов с другими нервными клетками. От возбуждения вставочных нейронов зависит, будет ли то или иное движение облегчено или заторможено. На спинальном уровне осуществляется наиболее простая форма автоматического регулирования состояния мышц – рефлекс на растяжение. В его основе лежит обратная связь от мышечных веретён к альфа-мотонейронам спинного мозга.
Управление работой мышц осуществляется не отдельными нейронами, а мотонейронным пулом, который определяет силу и участие в сокращении всех или части волокон каждой мышы. Мотонейронным или двигательным пулом называются мотонейроны, иннервирующие одну мышцу и рассеянные по нескольким сегментам спинного мозга.
Рассмотренные выше механизмы поддержания мышечного тонуса осуществляются на уровне спинного мозга, поэтому такой тонус называется спинальный. Он характеризуется очень слабой выраженностью тонического напряжения мышц и не может обеспечить поддержание позы животного и акт ходьбы. Но он достаточен для осуществления простейших спинальных рефлексов.
Сгибательные и ритмические рефлексы спинного мозга. Двигательные программы спинного мозга.
В моторных системах можно выделить три типа совершаемых движений: рефлекторные, ритмические и произвольные. Они различаются по степени сложности и участия сознания в контроле выполняемых действий. В каждом из них есть два компонента, нуждающиеся в контроле: тонический, который обеспечивает необходимую для начала движения позу и на время фиксирует в стабильном положении некоторые суставы, и фазный, определяющий направление и скорость движения. Рефлекторные движения относятся к самым простым моторным действиям, они выполняются быстро, стереотипно и в принципе не нуждаются в сознательном контроле, т.е. могут возникать непроизвольно. В качестве примера можно привести все сухожильные рефлексы (такие как коленный), а также защитный сгибательный рефлекс, возникающий в ответ на прикосновение к горячему предмету.
Сто человек из ста, нечаянно прикоснувшиеся рукой к сильно нагретому предмету, немедленно согнут её, что уберегает от ещё большего повреждения. Эта стереотипная защитная реакция возникает раньше, чем осознаётся смысл происшедшего. Она обеспечивается врожденным рефлекторным механизмом, в котором участвуют болевые чувствительные окончания, сенсорный нейрон, интеронейроны спинного мозга и мотонейроны мышц-сгибателей. По такому же рефлекторному стереотипу человек, наступивший босой ногой на колючку или острый камешек, немедленно ее сгибает. Это эволюционно древний рефлекс: ведь даже лишённая головного мозга лягушка сгибает лапку, погружённую в кислоту.
Некоторые двигательные рефлексы состоят из повторяющихся ритмических движений. Например, у многих шерстистых животных в ответ на неприятное раздражение кожи паразитами наблюдается почёсывание. Шеррингтон описал чесательный рефлекс у спинальных собак, что указывает на локализацию его центра в спинном мозгу. У спинальных животных были также выявлены локомоторные рефлексы, состоящие в ритмитческой смене сокращений сгибателей и разгибателей на разных лапах, как при ходьбе. И действительно, в спинном мозге обнаружена цепь нейронов, выполняющая функции генератора шагания. Она ответственна за чередование периодов возбуждения и торможения различных мотонейронов и может работать в автоматическом режиме.
Следовательно, простые двигательные стереотипные программы спинной мозг может осуществлять самостоятельно в зависимости от характера сенсорной информации. Он может, например, прервать сгибание пальцев, берущих какой-нибудь предмет, если этот предмет окажется сильно нагретым.
Запомните! СПИННОЙ МОЗГ
Мышечные веретёна расположены в толще скелетных мышц параллельно мышечным волокнам и прикреплены одним концом к мышце, другим - к сухожилию. Возбуждение в мышечном веретене возникает при удлинении мышечных волокон и сухожилий, т.е. при расслаблении или пассивном растяжении мышцы. Мышечные веретёна- рецепторы растяжения. В них при растяжении мышцы увеличивается частота нервных импульсов. При изотоническом сокращении мышцы частота импульсов снижается или прекращается. Сухожильные тельца Гольджи,наоборот, растягиваются и возбуждаются при мышечном сокращении. Импульсы от них по афферентным нервным волокнам поступают в ЦНС. Таким образом, мышечные веретёна регистрируют изменение длины мышцы, а рецепторы сухожилий - её напряжение (тонус).
Рефлексы растяжения
Импульсы от мышечных веретён при растяжении мышцы поступают к мотонейронам спинного мозга, в результате мышца сокращается. Такую рефлекторную дугу, в которую входит только один синапс, называют моносинаптической.Самый известный моносинаптический рефлекс - коленный. Эти рефлексы регулируют длину мышцы и особенно важны для мышц, поддерживающих позу при беге, ходьбе.
При сокращении мышц возбуждаются сухожильные рецепторы с одновременным торможением мотонейронов спинного мозга той же стороны. Ослабление мышечного тонуса активирует мотонейроны. Таким образом, рефлексы сухожильных рецепторов поддерживают постоянный мышечный тонус.
Реципрокное торможение
В регуляции движений принимают участие мышцы-сгибатели и разгибатели, состоящие в реципрокных (антагонистических) отношениях. От чувствительных нейронов спинномозговых ганглиев отходят волокна, которые ветвятся в спинном мозгу. Одни из них возбуждают нейроны, иннервирующие мышцы-сгибатели; а другие в то же самое время активируют вставочные нейроны, которые тормозят мотонейроны мышц-разгибателей.
ВИСЦЕРАЛЬНАЯ СЕНСОРНАЯ СИСТЕМА
Висцеральный анализатор
Анализатор внутренних органов (висцеральный, интероцептивный) участвует в поддержании постоянства внутренней среды организма (гомеостаза).
Интероцептивный анализатор имеет три отдела. Периферический отдел висцерального анализатора составляют интерорецепторы (висцерорецепторы),расположенные во внутренних органах. Проводниковый отдел представлен блуждающим, чревными, внутренностными тазовыми и другими нервами. Эфферентные вегетативные волокна этих нервов - симпатические и парасимпатические, а афферентные волокна - соматические. Центральный отдел висцерального анализатора расположен в моторной и премоторной областях коры больших полушарий.
Висцерорецепторы обладают низким порогом раздражения и высокой специфичностью к определённым раздражителям. Во внутренних органах есть рецепторы, реагирующие на изменения: давления (прессорецепторы), механические раздражения (механорецепторы), действие химических веществ (хеморецепторы), изменение температуры (терморецепторы), осмотического давления (осморецепторы). Висцерорецепторы участвуют в рефлекторных взаимодействиях внутренних органов, осуществляя висцеро-висцеральные и висцеро-кутанные рефлексы.
Висцеральная афферентация не всегда осознаваема. В целом она вызывает приятные или неприятные ощущения, определяемые как самочувствие и сильно влияющие на эмоциональное состояние. Кроме того, при раздражении глюкозных, осмотических рецепторов возникают вполне определенные чувства голода, жажды, а при раздражении рецепторов сфинктеров - позывы на мочеиспускание и дефекацию.
НОЦИЦЕПТИВНАЯ (БОЛЕВАЯ) СЕНСОРНАЯ СИСТЕМА
Значение болевых ощущений
Типы боли
Различают боль соматическую и висцеральную. Поверхностная соматическая больвозникает в коже, она бывает ранняя и поздняя. Боль в соединительной ткани, мышцах, суставах, костях называют глубокой; как правило, она тупая, трудно локализуемая и склонная к иррадиации. Висцеральная больможет быть вызвана быстрым растяжением органа (лоханки почек, стенки мочевого пузыря) или его спазмом (быстрым сокращением органа). Боль бывает острой и хронической. Острая боль(например, от ожога кожи) ограничена поврежденной областью, быстро исчезает после устранения повреждения. Устойчивые и рецидивирующие боли, длящиеся более полугода, называют хроническими(например, головные боли при мигрени, боли в сердце при стенокардии). Зудсчитают формой боли, возникающей в особых условиях.
Выделяют четыре компонента боли: сенсорный, эмоциональный, вегетативный и психомоторный. Вегетативный компонент- это реакция ВНС на болевую стимуляцию. При висцеральной боли (например, желчной колике) реакция ВНС может быть очень сильной и проявляться тошнотой, рвотой, потоотделением, падением АД. Двигательный компонентболи проявляется рефлексом избегания или защиты (например, отдергивание руки от горячего). При висцеральной боли этот рефлекс приобретает форму мышечного напряжения. Обычно все компоненты боли возникают вместе, но проявляются по-разному.
Адаптации к боли, видимо, не происходит (головная и зубная боль может длиться часами, причиняя страдание), однако возможно привыкание к многократным болевым стимулам.
Ноцицептивные рецепторы
В настоящее время считают, что боль - это специфическое ощущение с собственным специализированным нервным аппаратом: рецепторами, проводящими путями и нервными центрами.
Рецепторы боли - ноцирецепторы- это свободные нервные окончания, расположенные в глубоких слоях эпидермиса, в сосочковом слое дермы, в адвентициальной оболочке мелких кровеносных и лимфатических сосудов, в соединительной ткани мышц, суставов и сухожилий. В коже они не рассеяны диффузно, а сосредоточены в болевых точках.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Необходимым условием нормальной мышечной деятельности является получение информации о положении тела в пространстве и о степени сокращения каждой из мышц. Эта информация поступает в центральную нервную систему от рецепторов вестибулярного аппарата, глаз, кожи, а также от проприорецепторов (мышечно-суставных рецепторов). К проприорецепторам относятся:
- мышечные веретена, находящиеся среди мышечных волокон,
- тельца Гольджи, расположенные в сухожилиях,
- пачиниевы тельца, находящиеся в фасциях, покрывающих мышцы, в сухожилиях, связках и периосте.
Все эти проприорецепторы относятся к группе механорецепторов. Мышечные веретена и тельца Гольджи возбуждаются при растяжении, а пачиниевы тельца — при давлении.
О важности афферентной иннервации мышц свидетельствует и тот факт, что 30-50% волокон в любом нерве, подходящем к мышце, являются афферентными, несущими импульсацию от проприорецепторов. К уже отмечалось, при выключении афферентной иннервации мышц, например при перерезке задних корешков спинного мозга, координированная мышечная деятельность нарушается, несмотря на сохранение двигательной иннервации мышц.
Мышечные веретена представляют собой высокодифференцированные рецепторные образования, снабженные афферентными и эфферентными нервными волокнами. Каждое веретено состоит из нескольких тонких так называемых интрафузальных поперечно-полосатых мышечных волокон.
Oдиночное волокно состоит из центральной части — ядерной сумки и двух способных к сокращению участков. В ядерной сумке pacположены рецепторы, представляющие сообой спиралевидные окончания толстых афферентных покрытых миелиновой оболочкой нервных волокон. Сокращающиеся участки интрафузального волокна иннервированы тонкими моторными, так называемыми гамма-эфферентными, волокнами, образующими мелкие концевые пластинки.
Один конец интрафузального волокна прикрепляется к обычному мышечному волокну, а другой — к сухожилию. Таким образом, интрафузальные волокна расположены параллельно массе мышечных волокон, среди которых они находятся. Если мышца растянута или расслаблена, то мышечные веретена тоже растянуты и в рецепторах ядерной сумки возникают импульсы, идущие в центральную нервную систему. Если же мышца сокращена, то натяжение мышечных веретен ослабевает и импульсация прекращается.
Второй вид проприорецепторов — тельца Гольджи — находится в сухожилиях. При мышечном сокращении тельца Гольджи растягиваются и в них возникают нервные импульсы, поступающие к нервным центрам. Сухожильные рецепторы менее возбудимы, чем мышечные веретена.
Таким образом, в двигательном аппарате имеются рецепторы — мышечные веретена,— возбуждающиеся при удлинении (расслаблении и растяжении), и рецепторы — тельца Гольджи,— возбуждающиеся при сокращении мышечных волокон.
Импульсы, поступающие в центральную нервную систему от мышечных веретен, облегчают возникновение рефлекторной реакции данной мышцы и тормозят сокращение мышцы-антагониста. Импульсы, приходящие от сухожильных рецепторов — телец Гольджи, вызывают противоположные рефлекторные реакции.
Импульсация из мышечных веретен, как показал Р. Гранит, может изменяться с помощью специального регуляторного механизма. Такой механизм образован сократительными элементами, расположенными по обе стороны от ядерной сумки интрафузального волокна. Их сокращение вызывает растяжение ядерной сумки и раздражение рецепторов веретена. Таким образом, эти рецепторы могут посылать в центральную нервную систему интенсивную импульсацию даже при небольшом сокращении или расслаблении поперечнополосатой мышцы.
Степень сокращения сократительных элементов веретена регулируется гамма-эфферентными нервными волокнами, которые являются отростками, гамма-мотонейронов спинного мозга. Импульсы, приходящие по гамма-эфферентным волокнам, вызывают сокращения интрафузальных мышечных волокон, что приводит к усилению потока афферентных импульсов от растягиваемых рецепторов ядерной сумки. Активность гамма-мотонейронов регулируется ретикулярной формацией среднего мозга.
Сократительные элементы мышечных веретен всегда находятся в состоянии некоторого тонуса, так как по гамма-эфферентам непрерывно поступают к ним импульсы из центральной нервной системы. Это влечет собой афферентную импульсацию от мышечных веретен, которая в свою ередь тонизирует мотонейроны спинного мозга, что является одной из причин рефлекторного тонуса скелетной мускулатуры. Отличительная особенность проприорецепторов — их малая способность к адаптации, благодаря чему центральная нервная система получает непрерывные сигналы о состоянии скелетной мускулатуры и может осуществлять непрерывную регуляцию двигательных актов.
Кинестетические сигналы, т. е. сигналы о движении отдельной части тела, играют чрезвычайно важную роль в развитии восприятий организма. Они являются главным контролем других органов чувств, например, зрения. Так, оценка зрением расстояния до какого-нибудь предмета вырабатывается при помощи мышечного чувства. Возбуждения, связанные с работой мышц, возникающие при хватании предмета руками или приближении к нему, сочетаются со зрительными ощущениями и служат важнейшим критерием для выработки суждения о том, насколько отдален предмет.
В мышцах человека содержатся три типа специализированных рецепторов: первичные окончания веретен, вторичные окончания веретен и сухожильные рецепторы Гольджи. Эти рецепторы реагируют на механические раздражения и участвуют в координации движений, являясь источником информации о состоянии двигательного аппарата.
Мышечные веретена представляют собой небольшие продолговатые образования, расположенные в толще мышцы (рис.). Внутри капсулы каждого веретена находится пучок мышечных волокон. Эти волокна называют интрафузальными, в отличие от всех остальных волокон мышцы, которые носят название зкстрафузальные. Веретена расположены параллельно экстра-фузальным волокнам, поэтому при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении уменьшается.

Рис. Строение мышечного волокна и его иннервация.
1 — проксимальный конец прикрепленной к волокну скелетной мышцы; 2 — дистальный конец этого волокна, прикрепленного к фасции; 3 — ядерная сумка; 4 — афферентные волокна; 5 — эфферентные волокна; 6 — моторное волокно, идущее к скелетной мышце.
Веретена реагируют на два типа воздействии: изменение длины мышцы и изменение уровня активации системы эфферентных волокон. При растяжении пассивной мышцы наблюдается активация рецепторов (мышечных веретен).
Веретена можно рассматривать как непосредственный источник информации о длине мышцы, о ее изменениях, если только мышца не возбуждена. При активном состоянии мышцы необходимо учитывать влияние системы, которая поддерживает импульсацию веретен укорачивающейся мышцы, что дает возможность рецепторам реагировать на вызванные помехами неравномерности движения как увеличением, так и уменьшением частоты импульсов и участвовать таким образом в коррекции движений.
Разминая мышцу, массажист добивается усиления импульсации с мышечных веретен в соответствующие сегменты спинного мозга, тем самым повышая тонус того нервного центра, который управляет деятельностью этой мышцы.
Возникает вопрос: почему, несмотря на то, что применение любых массажных приемов вызывает усиление импульсации с рецепторов мышечных веретен или механорецепторов и — как следствие — повышение тонуса соответствующих отделов центральной нервной системы, кроме возбуждающего действия массажа, различают и его тормозные эффекты? Этот феномен можно объяснить одним из свойств нервных центров, куда поступает вся информация о внешних воздействиях на организм и о его внутреннем состоянии.
Безусловно, следует иметь в виду, что структура и характер ответных реакций организма на массаж каждый раз бывают различными в зависимости от соотношения процессов возбуждения и торможения в коре головного мозга, функционального состояния рецепторного поля, которое подвергается массажному воздействию, состояния тренированности, характера массажа и используемых приемов. Эффективность массажа зависит от знания массажистом функционального состояния массируемого.
К группе проприорецепторов, кроме мышечных веретен, относятся и суставные рецепторы. Они располагаются в суставных сумках и в связочном аппарате.
В суставных сумках находятся рецепторы, у которых частота разрядов изменяется в соответствии как с положением сустава в покое, так и со скоростью его движения. Гистологически они представляют собой рецепторы типа телец Руффини. Связки содержат рецепторы, сходные по форме с тельцами Гольджи, и немного телец, подобных тельцам Пачини. Кроме того, имеются свободные нервные окончания, функция которых пока неизвестна. Предполагают, что они участвуют в передаче болевых сигналов от суставов.
При выполнении массажистом пассивных движений в суставах у массируемого возбуждаются суставные рецепторы и инициируется цепь процессов, сходных по своим физиологическим механизмам с рассмотренными выше особенностями передачи нервных импульсов от мышечных веретен и механо-рецепторов.
В заключение, посвященное физиологическому воздействию массажа на механо- и проприорецепторы, следует отметить, что в них влияние массажа рассматривалось только с одной стороны — с точки зрения нервно-рефлекторной теории. Однако, как уже указывалось ранее, эффекты, получаемые при применении массажных приемов, могут быть объяснены только при учете комплексного воздействия массажа на организм. Для получения более полной картины физиологического воздействия массажа необходимо рассмотреть другие стороны его влияния.
Проприоцепция — это группа сигналов, посылаемых в центральную нервную систему специальными терминалами (проприоцепторами), расположенными в суставных капсулах, связках, сухожилиях и мышцах.
Проприоцепторы, среди которых выделяют мышечные рецептор ных образований" href="http://www.braintools.ru/article/9580"> рецептор ы, или мышечные веретена ( рецептор ы растяжения), сухожильные рецептор ы, или органы Гольджи ( рецептор ы мышечной силы), а также суставные рецепторы относятся к механо рецептор ам, посылающим в ЦНС информацию о положении, деформации и смещениях различных частей тела.
Функционирование этих рецептор ов обеспечивает координацию всех подвижных органов и тканей животного и человека в состоянии покоя и во время любых двигательных актов. При экспериментальном выключении проприоцепторов животные теряют способность поддерживать естественные позы, двигаться и целесообразно реагировать на внешние воздействия.
Если человек закроет глаза и попытается написать текст, то буквы будут написаны достаточно четко. Этим простым способом легко убедиться в умении человека пользоваться информацией, идущей от мышц и суставов.
Проприоцепторы составляют периферическую часть проприоцептивной сенсорной системы , или двигательного анализатора . Вместе с тем, несмотря на то, что миллионы людей ежедневно пользуются услугами этого анализатора, мы до сегодняшнего дня знаем сравнительно мало о его деятельности. Это особенно касается работы коркового отдела двигательного анализатора. Внутренние проприоцепторы находятся в мышцах, сухожилиях, сухожильных влагалищах, межкостных мембранах, фасциях, тканях суставов, надкостнице и т.д. Среди них имеются неспециализированные рецептор ы, встречающиеся и в других частях тела (свободные нервные окончания, инкапсулированные рецептор ы типа телец Руффини и Пачини), и специализированные — мышечные веретена и сухожильные органы (или рецептор ы) Гольджи.
Двигательный, или кинестетический, анализатор (мышечная сенсорная система ) обеспечивает формирование так называемого мышечного чувства при изменении напряжения мышц, суставных сумок, связок и сухожилий. Проблема мышечно-суставных ощущений имеет исключительное значение для физиологии и психологии. Специфические особенности человека появились благодаря более совершенной организации мышечной сенсорной системы по сравнению с животными.
И.М. Сеченов считал, что мышечное чувство является ближайшим регулятором движений и одним из орудий ориентации человека в пространстве и времени. Ощущения положения и перемещения тела в пространстве, ощущения во время трудовой деятельности и членораздельная речь лежат в основе формирования сознания человека и его представлений об окружающем мире.
Мышечное чувство обладает тремя качествами. Это, во-первых, ощущение положения конечностей, когда человек может определить положение своих конечностей и их частей относительно друг друга. Во-вторых, ощущение движения, когда, изменяя угол сгибания в суставе, человек осознаёт скорость и направление движения. Третьим качеством является ощущение усилия, когда человек может оценить мышечную силу, нужную для движения или удерживания суставов в определённом положении при подъёме или перемещении груза.
Наряду с кожной, зрительной и вестибулярной сенсорными системами двигательный анализатор оценивает положение тела в пространстве, позу, участвует в координации мышечной деятельности.
Двигательная сенсорная система служит для анализа состояния двигательного аппарата — его движения и положения. Информация о степени сокращения скелетных мышц, натяжении сухожилий, изменении суставных углов необходима для регуляции двигательных актов и поз.
Общий план организации. Двигательная сенсорная система состоит из следующих 3-х отделов:
- периферический отдел, представленный проприо рецептор ами, расположенными в мышцах, сухожилиях и суставных сумках;
- проводниковый и отдел, который начинается биполярными клетками (первыми нейрон ами), тела которых расположены вне ЦНС — в спинномозговых узлах. Один их отросток связан с рецептор ами, другой входит в спинной мозги передает проприоцептивные импульсы ко вторым нейрон ам в продолговатый мозг (часть путей от проприо рецептор ов направляется в кору мозжечка ), а далее к третьим нейрон ам — релейным ядрам таламуса (в промежуточный мозг);
- корковый отдел находится в передней центральной извилине коры больших полушарий.
Функции проприо рецептор ов
К проприо рецептор ам относятся мышечные веретена, сухожильные органы (или органы Гольджи) и суставные рецептор ы ( рецептор ы суставной капсулы и суставных связок). Все эти рецептор ы представляют собой механо рецептор ы, специфическим раздражителем которых является их растяжение.
Мышечные веретена человека, представляют собой продолговатые образования длиной несколько миллиметров, шириной десятые доли миллиметра, которые расположены в толще мышцы. В разных скелетных мышцах число веретен на1 г ткани варьирует от нескольких единиц до сотни.
Таким образом, мышечные веретена как датчики состояния силы мышцы и скорости ее растяжения реагируют на два воздействия: периферическое — изменение длины мышцы, и центральное — изменение уровня активации гамма-мото нейрон ов. Поэтому реакции веретен в условиях естественной деятельности мышц довольно сложны. При растяжении пассивной мышцы наблюдается активация рецептор ов веретен; она вызывает миотатичес-кий рефлекс , или теор ии рефлекторной деятельности" href="http://www.braintools.ru/article/8998">рефлекс на растяжение. При активном сокращении мышцы уменьшение ее длины оказывает на рецептор ы веретена дезактивирующее действие, а возбуждение гамма-мото нейрон ов, сопутствующее возбуждению альфа-мото нейрон ов, приводит к реактивации рецептор ов. Вследствие этого импульсация от рецептор ов веретен во время движения зависит от длины мышцы, скорости ее укорочения и силы сокращения.
Сухожильные органы ( рецептор ы) Гольджи человека располагаются в зоне соединения мышечных волокон с сухожилием, последовательно по отношению к мышечным волокнам.
Сухожильные органы представляют собой структуру вытянутой веретенообразной или цилиндрической формы, длина которой у человека может достигать1 мм. Этот первичночувствующий рецептор . В условиях покоя, т.е. когда мышца не сокращена, от сухожильного органа идет фоновая импульсация. В условиях мышечного сокращения частота импульсации возрастает прямо пропорционально величине мышечного сокращения, что позволяет рассматривать сухожильный орган как источник информации о силе, развиваемой мышцей. В тоже время сухожильный орган слабо реагирует на растяжение мышцы.
В результате последовательного крепления сухожильных органов к мышечным волокнам (а в ряде случаев — к мышечным веретенам), растяжение сухожильных механо рецептор ов происходит при напряжении мышц. Таким образом, в отличие от мышечных веретен, сухожильные рецептор ы информируют нервные центры о степени напряжения мыши, и скорости его развития.
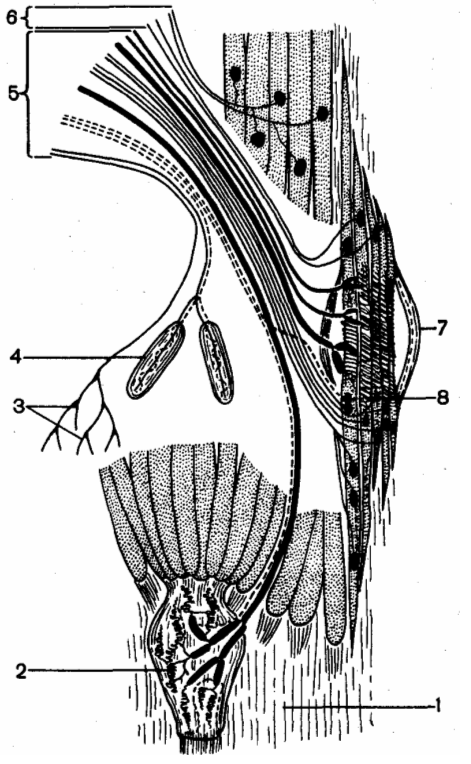
Эфферентная иннервация скелетной мышцы: 1 — сухожилие мышцы; 2 — сухожильные рецептор ы Гольджи; 3 — свободные окончания; 4 — тельца Фатера-Пачини; 5 — афферентные волокна; 6 — эфферентные волокна; 7 — мышечное веретено; 8 — окончания афферентных волокон
Суставные рецептор ы реагируют на положение сустава и на изменения суставного угла, участвуя, таким образом, в системе обратных связей от двигательного аппарата и в управлении им. Суставные рецептор ы информируют о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецептор ы представляют собой свободные нервные окончания или окончания, заключенные в специальную капсулу. Одни суставные рецептор ы посылают информацию о величине суставного угла, т. е. о положении сустава. Их импульсация продолжается в течение всего периода сохранения данного угла. Она тем большей частоты, чем больше сдвиг угла. Другие суставные рецептор ы возбуждаются только в момент движения в суставе, т. е. посылают информацию о скорости движения. Частота их импульсации возрастаете увеличением скорости изменения суставного угла.
Проводниковый и корковый отделы проприоцеп-тивного анализатора млекопитающих и человека. Информация от мышечных, сухожильных и суставных рецептор ов поступает через аксон ы первых афферентных нейрон ов, находящихся в спинномозговых ганглиях, в спинной мозг , где частично переключается на альфа-мото нейрон ы или на вставочные нейрон ы (например, на клетки Реншоу), а частично направляется по восходящим путям в высшие отделы головного мозга. В частности, по путям Флексига и Говерса проприоцептивная импульсация доставляется к мозжечку, а по пучкам Голля и Бурдаха, проходящим в дорсальных канатиках спинного мозга , она доходит до нейрон ов одноименных ядер, расположенных в продолговатом мозге.
Аксоны таламических нейрон ов ( нейрон ов третьего порядка) оканчиваются в коре больших полушарий, главным образом, в соматосенсорной коре (постцентральная извилина) и в области сильвие-вой борозды (соответственно, участки S-1 и S-2), а также частично в двигательной (префронтальной) области коры. Эта информация используется двигательными системами мозга достаточно широко, в том числе для принятия решения о замысле движения, а также для его реализации. Кроме того, у человека на основе про-приоцептивной информации формируются представления о состоянии мышц и суставов, а также, в целом, о положении тела в пространстве.
Сигналы, идущие от рецептор ов мышечных веретен, сухожильных органов, суставных сумок и тактильных рецептор ов кожи. Механо рецептор ная и температурная чувствительность" href="http://www.braintools.ru/article/9885"> рецептор ов кожи, называют кинестетическими, т. е. информирующими о движении тела. Их участие в произвольной регуляции движений различно. Сигналы от суставных рецептор ов вызывают заметную реакцию в коре больших полушарий и хорошо осознаются. Благодаря им человек лучше воспринимает различия при движениях в суставах, чем различия в степени напряжения мышц при статических положениях или поддержании веса. Сигналы же от других проприо рецептор ов, поступающие преимущественно в мозжечок, обеспечивают бессознательную регуляцию, подсознательный контроль движений и поз.
Таким, образом, проприоцептивные ощущения дают человеку возможность воспринимать изменения положения отдельных частей тела в покое и во время совершаемых движений. Информация, поступающая от проприоцепторов, позволяет ему постоянно контролировать позу и точность произвольных движений, дозировать силу мышечных сокращений при противодействии внешнему сопротивлению, например при подъеме или перемещении груза.
Проприоцептивное восприятие положения тела и движений происходит в результате объединения в соматосенсорной коре информации от всех разновидностей проприоцепторов.
Читайте также:
