
Плазматические отростки мышечных клеток это
Известно более 12 000 видов нематод, обитающих в морях, пресных водах, почве и живых организмах. Длина тела круглых червей варьирует от до 8 метров.
Рассмотрим внешнее и внутреннее строение тела нематод на примере лошадиной аскариды.
Лошадиная аскарида (Parascaris equorum) ведёт паразитический образ жизни в тонком кишечнике лошади, питаясь продуктами расщепления и тельных веществ. Этот гельминт достигает длины до 37 – 40 см. Самки длинее самцов, хвостовой конец у последних загнут на брюшную сторону.
Тело аскариды веретеновидной формы, круглое на поперечном разрезе (Рис. 5). Передняя и задняя части тела сужены. Снаружи тело покрыто кутикулой, напоминающей по составу коллаген, и, поэтому, очень эластичной. Образование кутикулы происходит в результате деятельности клеток гиподермы (видоизменённый однослойный эпителий). Кутикула состоит из нескольких слоев клеток, причём нижний слой имеет волокнистое синцитиальное строение, Под гиподермой лежит базальная мембрана – неклеточная структура, отграничивающая эпителиальный слой от мышечного. Вдоль тела у большинства нематод проходят 4 продольные линии: две по бокам и по одной на спинной брюшной сторонах. На внутренней поверхности гиподерма образует утолщения в виде 4-х валиков, соответствующих продольным линиям. В боковых валиках гиподермы проходят каналы выделительной системы, а в спинном и брюшном – главные нервные стволы.Под базальной мембраной залегает слой продольных мышц. Каждая мышечная клетка состоит из плазматического отростка, саркоплазматического мешка (собственно тело клетки) и сократимой части. Валиками гиподермы мышечный слой разбит на 4 продольные ленты. Плазматические отростки мышечных клеток направлены либо в сторону спинного, либо брюшного валика гиподермы.
Рис. 5. Поперечный разрез тела самки аскариды
1- кутикула, 2- гиподерма, 3- продольные мышцы, 4- плазматические отростки мышечных клеток, 5- спинной валик гиподермы, 6- спинной нервный ствол, 7- боковой валик гиподермы, 8- выделительный канал, 9-матка, 10-яичник, 11-брюшной валик гиподермы, 12-брюшной нервный ствол, 13-яйцевод, 14-кишечник, 15-первичная полость тела.
Поэтому принято считать, что мышечный слой делится на спинные и брюшные мышцы-антагонисты. Кутикула, гиподерма и слой продольных мышц составляют кожно-мускульный мешок нематод. Внутри него образуется первичная полость тела, где располагаются органы пищеварительной и половой систем. Полость тела круглых червей называется первичной, т.к. сформировалась путём редукции паренхимы. Внутри первичная полость заполнена жидким содержимым. Полостная жидкость содержит органические кислоты (масляную, валериановую, капроновую и др.), образующиеся в результате анаэробного расщепления гликогена. Первичнополостная жидкость создаёт в теле аскариды избыточное давление и если по каким-то причинам внутри тела хозяина происходит разрыв стенки тела аскариды, то вылившаяся полостная жидкость гельминта вызывает мучительные боли.
Пищеварительная система аскариды состоит из передней, средней и задней кишки. Передняя кишка начинается ртом, находящимся на переднем конце тела и окружённым тремя губами. Рот ведёт в глотку. Глотка переходит в передний отдел средней кишки, часто называемый пищеводом, в стенках которого лежат пищеводные железы. У некоторых нематод, например, остриц, задняя часть пищевода образует колбообразное расширение – бульбус, являющийся систематическим признаком подотряда оксиурат (остриц). Средняя кишка выстлана цилиндрическим эпителием с ворсинками на внутренней поверхности. В протоплазме клеток эпителия кишечника откладываются запасные питательные вещества в виде зёрен гликогена. Содержание последнего в высушенном состоянии достигает 32%. Задняя кишка, как и передняя, выстлана изнутри кутикулой и заканчивается анальным отверстием, куда у самцов открывается и отверстие половой системы. Дыхательная система отсутствует. Дыхание осуществляется анаэробным путем за счёт расщепления питательных веществ. Некоторые авторы приводят данные о наличии у аскарид дыхательного пигмента, подобного на гемоглобин млекопитающих.
Нервная система состоит из окологлоточного нервного кольца и отходящих от него нервных стволов. Нервное кольцо окружает переднюю часть пищевода и отдаёт к переди 6 коротких нервных стволов и к зади 6 длинных нервных стволов, из которых спинной и брюшной наиболее развиты и проходят в валиках гиподермы. Оба ствола соединяются между собой в виде полуколец многочисленными перемычками – комиссурами.
Органы чувств нематод развиты слабо. Вокруг рта, а у самцов на заднем конце тела располагаются осязательные бугорки и щетинки. По бокам головного конца тела лежат амфиды – органы химического чувства.
Выделительная система весьма своеобразного строения. В боковых валиках гиподермы проходят выделительные каналы, начинающиеся слепо в задней части тела и сливающиеся в срединный непарный канал, который открывается выводным протоком позади губ. По гистологическим данным, эти каналы представляют собой одну гигантскую клетку (т.н. шейную железу) с ядром, лежащим на уровне передней части пищевода. Ядро этой клетки носит название синкариона. На боковых валиках гиподермы лежат две пары фагоцитарных клеток. Из полостной жидкости эти клетки поглощают продукты распада и инородные тела. Нерастворимые частицы, накапливаются в фагоцитарных клетках и никуда не удаляются из них на протяжении всей жизни аскариды.
В половом отношении большинство нематод раздельнополы. Половая система самки лошадиной аскариды состоит из тонких, нитевидных парных яичников, которые, расширяясь, переходят в яйцеводы, где происходит формирование яиц. Яйцеводы без видимых границ переходят в парную матку, стволы которой сливаясь, образуют непарное влагалище, открывающееся половым отверстием на брюшной стороне тела. У самца имеется единственный нитевидный семенник, переходящий без резких границ в канал большего диаметра – семяпровод. Под кишечником он направляется кзади и образует семенной пузырь, где накапливается сперма. Пузырь сужается в тонкую мускулистую трубку – семтвергательный канал, впадающий в заднюю кишку. Туда же открывается парная совокупительная сумка (половая бурса), в которой помещаются две кутщлярные иглы, или спикулы. Они высовываются через анальное отверстие наружу и служат в качестве вспомогательных органов при совокуплении.
Выделенные во внешнюю среду яйца параскарид при благоприятных условиях (температуре 18 – 20 °С и влажности не менее 60%) становятся инвазионными в течение 7 – 14 дней. Из яиц в кишечнике лошади выходят личинки, куда яйца попадают вместе с кормом или водой. Через слизистую оболочку кишечника личинки проникают в кровеносные сосуды и заносятся током крови в легкие. В легочных альвеолах личинки растут, линяют и мигрируют в бронхи. Это вызывает раздражение дыхательных путей и рефлекторный кашель. Вместе с мокротой личинки параскарид попадают в ротовую полость и заглатываются. В кишечнике они развиваются до половозрелой стадии и приступают к размножению. Срок развития параскарид до половой зрелости составляет около 2 – 2,5 месяцев. В цикле развития лошадиной аскариды отсутствует промежуточный хозяин, поэтому её относят к группе геогельминтов. Аналогичным образом происходит развитие человеческой и свиной аскарид.
Цикл развития лошадиной аскариды был рассмотрен выше, поэтому остановимся на циклах развития следующих паразитических нематод: свиной и человеческой аскарид, токсокар и токсаскарид, власоглава, детской острицы, метастронгилюса и трихинеллы.
Свиная аскарида (Ascaris suis) паразитирует в тонком кишечнике домашних и диких свиней. Цикл развития проходит без участия промежуточного хозяина, т.е. свиная аскарида относится к геогельминтам. За сутки одна половозрелая самка аскариды откладывает до 200 000 яиц, которые с фекалиями попадают во внешнюю среду. Там, при благоприятных условиях (температура 20 – 30 С и влажность более 60 %), через 2 – 3 недели внутри яиц развиваются инвазионные личинки. Свиньи заражаются аскаридами алиментарным путём. В кишечнике свиньи личинки выходят из яиц и внедряются в слизистую оболочку, проникают в капилляры и током крови заносятся в печень, а оттуда – в лёгкие. В лёгких личинки аскарид по мере развития проникают в альвеолы, бронхиолы, бронхи и трахею. Раздражая рецепторы слизистой оболочки дыхательной системы, личинки вызывают рефлекторный кашель. При откашливании, с мокротой, они заносятся в ротовую полость и вновь поступают в тонкий кишечник. Гам, через 1,5 – 2,5 месяца личинки аскарид достигают половой зрелости и приступают к размножению.
Человеческая аскарида (Ascaris lumbricoides) вызывает аскариоз человека, паразитируя в тонком отделе кишечника. Цикл развития человеческой аскариды во многом аналогичен таковому свиной аскариды. Наиболее тяжело аскаридоз протекает у детей, известны случаи непроходимости кишечника из-за наличия в нём большого количества паразитов. Профилактика аскаридоза сводится к соблюдению гигиенических правил, а лечение – к применению лекарственных препаратов (антгельминтиков), наименее токсичным из которых является пиперазин.
Власоглав (Trichocephalus trichiuris) паразитирует в толстом отделе кишечника человека. Название паразита связано с тем, что передняя часть тела гельминта по форме напоминает волос. Задняя часть тела, наоборот резко утолщена. Головным концом власоглав внедряется в слизистую оболочку кишечника, а его задняя часть находится в просвете кишки. Питается власоглав кровью хозяина. Заражение человека трихоцефалёзом происходит алиментарным путём. Власоглав относится к геогельминтам. Попавшие во внешнюю яйца становятся инвазионными при благоприятных условиях через 1 – 1,5 месяца. Лечение при трихоцефалёзе затруднительно в связи с прочной фиксацией власоглавов в слизистой оболочке кишечника. Домашние и дикие свиньи также подвержены трихоцефалёзу, вызываемому Trichocephalus suis. Кроме свиней, власоглавы паразитируют у жвачных и плотоядных.
Детская острица (Enterobius vermicularis) (Рис. 6) достигает длины до1см.
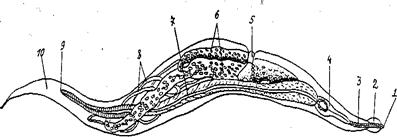
Рис. 6. Самка детской острицы
1 - рот; 2 - кутикулярные утолщения; 3 - пищевод; 4 - бульбус; 5 - половое отверстие; 6 - матка, заполненная яйцами; 7 - средняя кишка; 8 - яичник; 9 - анальное отверстие; 10 – хвост
Задний конец тела паразита заострен. Паразитирует в толстом отделе кишечника человека, чаще у детей. Заболевание, вызываемое детской острицей, называется энтеробиоз. В вечернее и ночное время самки остриц выходят анальное отверстие и откладывают яйца на кожу в области промежности, вызывает зуд. Ребёнок расчёсывает зудящие места руками и яйца остриц попадают в подногтевые пространства, а оттуда – в ротовую полость, и далее, в кишечник. Там личинки выходят из яиц, достигают половой зрелости и размножаются. Острицы вызывают воспаление слизистой оболочки толстого отдела кишечника и могут явиться причиной аппендицита. Питаются они содержимым кишечника. Переносчиками яиц остриц могут служить мухи, тараканы и другие насекомые.
Трихинелла (Trichinella spiralis) вызывает заболевание диких и домашних животных, а также человека, под названием трихинеллёз. Сами нематоды очень мелкие, до 1,6 – 4 мм длиной, самки крупнее самцов. Особенностью биологии 1рихинелл является то, что они живородящие, развиваются только в организме хозяев без выхода во внешнюю среду. При этом, одно животное может выступать одновременно в роли промежуточного и дефинитивного хозяина. Заражение трихинеллёзом происходит при поедании трихинеллёзного мяса, в котором содержатся живые личинки паразита. После попадания в кишечник они становятся половозрелыми через 30 – 48 часов. Самки трихинелл внедряются в слизистую оболочку кишечника, оплодотворяются самцами, которые после этого погибают, и через четверо суток рождают живых личинок. Они гематогенным путём заносятся в поперечнополосатые мышцы, где растут, спирально закручиваясь, Примерно через месяц вокруг личинок образуются соединительнотканные капсулы, которые подвергаются обызвествлению через 6 – 16 месяцев. В сердечной поперечнополосатой мышечной ткани личинки трихинелл не развиваются. Инкапсулированные личинки в мышцах могут сохранять жизнеспособность более 25 лет. Заражение человека трихинеллами происходит при употреблении в пищу трихинеллёзного мяса свиней и диких животных. Плотоядные и грызуны являются природным резервуаром трихинеллёза. В качестве профилактики этого опасного заболевания необходимо проводить ветеринарно-санитарную экспертизу туш свиней и диких животных. При содержании свиней следить, чтобы они не поедали дохлых крыс, мышей и боенские отходы. Заражённые трихинеллами туши животных необходимо уничтожать путем автоклавирования или сжигания. В Беларуси периодически отмечаются случаи заболевания людей трихинеллёзом, особенно среди охотников.
Аскарида человеческая (Ascaris lumbricoides). Тело аскариды несегментированное, цилиндрической формы, заостряющееся с обоих концов.
Живая аскарида имеет красновато-желтый цвет, фиксированная — беловатый. Аскариды — раздельнополые, причем самки значительно больше самцов: длина самок 20 — 30 см, длина самцов 15 — 20 см. Самки и самцы отличаются друг от друга не только по размерам; задний конец самцов изогнут или даже закручен на брюшную сторону. Передний конец тела у обоих полов одинаков: он несколько тоньше заднего и несет на себе ротовое отверстие, окруженное тремя небольшими вздутиями — губами.
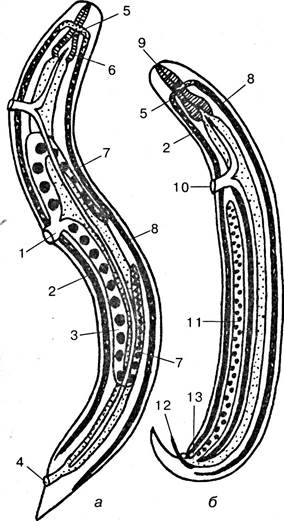
У самцов брюшной стороной считается та, на которую загнут задний конец тела, у самки на брюшной стороне ближе к переднему концу тела лежит половое отверстие, а ближе к заднему концу — анальное отверстие (рис. 3.47).
Рис. 3.47. Строение нематод: а — самка; б — самец: 1— влагалище; 2 — вентральный нервный тяж; 3 — матка; 4 — анус; 5 — нервное кольцо; б — глотка; 7— яичник; 8 — дорзальный нервный тяж; 9 — рот; 10 — выделительный канал; 11 — семенник; 12 — спикула; 13 — клоака
Все тело аскариды покрыто блестящей белой кутикулой и лишено ресничного покрова.
Покров. Кожно-мускульный мешок состоит из трех слоев: кутикулы, гиподермы и продольной мускулатуры.
Кутикула имеет многочисленное строение и выполняет функцию защиты и наружного скелета, к которому прикрепляются мышцы.
Гиподерма представлена симпластической тканью с беспорядочно расположенными ядрами. У большинства нематод всегда имеется четыре продольных валика, вдающихся в полость тела.
Мускулатура представлена одним слоем продольных мышц, разделенных валиком гиподермы на четыре продольные ленты. Каждая мышечная клетка состоит из сократимой части, представленной веретеном, и плазматического выроста (плазматическая часть). Движение ограничено. Тело изгибается только в дорзовентральной плоскости благодаря тому, что брюшные и спинные продольные мышечные ленты действуют как антагонисты.
Полость тела. Первичная или псевдоцель. Она охвачена мышечной тканью и не имеет эпителиальной выстилки. Псевдоцель содержит полостную жидкость и клеточные элементы. Полостная жидкость находится под большим давлением, что создает опору для мышечного слоя (гидроскелет). Она играет важную роль в обменных процессах. Обладает своеобразным запахом, который зависит от присутствия в этой жидкости валериановой и других кислот. В псевдоцеле находятся внутренние органы (рис. 3.48).
Рис. 3.48. Поперечный разрез аскариды: 1— кутикула; 2 — гиподерма; 3 — боковой валик с каналом выделительной системы; 4 — первичная полость тела; 5 — плазматические отростки мышечных клеток; 6 — мускулатура; 7 — брюшной нервный ствол; 8 — матка; 9 — яйцеводы; 10 — яичник; 11 — кишечник; 12 — спинной нервный ствол

Нервная система. Основной ее частью является окологлоточное нервное кольцо. Оно охватывает пищевод и отсылает в разные стороны целый ряд стволов. Крупнейшие из них — брюшной и спинной стволы — тянутся вдоль брюшного и спинного гиподермальных валиков. По бокам тела имеются два боковых (латеральных) нервных ствола. Наконец, у аскариды имеются еще две пары продольных стволов — два субдорзальных и два субвентральных.
От нервного кольца кпереди направляется ряд нервов, иннервирующих головной конец. Кроме того, от нервных стволов идут комиссуры, соединяющие нервные стволы между собой.
Нервная система полностью находится в кожно-мускульном мешке.
Органы чувств. Специальных органов чувств у нематод обычно нет. Кожа их отличается малой чувствительностью и слабовыраженной раздражимостью. В ней встречаются лишь отдельные нервные окончания, чувствительные щетинки и сосочки, особенно в головном и анальном концах тела, являющиеся рецепторами химических чувств и осязания.
Иногда у свободноживущих нематод имеются пигментированные глазки, даже с линзой.
Пищеварительная система представлена трубкой, кото рая состоит из передней (эктодермальной), средней (эндодермальной) и задней (экдодермальной) кишок.
Передняя кишка начинается ротовым отверстием, которое вооружено тремя губами. Эти губы играют роль присасывательного органа. Ротовое отверстие ведет в глотку и пищевод.
Пищевод имеет трехгранный просвет, выстланный кутикулой. Стенки пищевода обладают радиально расположенной мускулатурой, между глоткой и пищеводом имеется промежуточное вздутие — бульбус.
Средняя кишка — в виде прямой трубки, выстлана однорядным эпителием. Поверхность эпителия покрыта ворсинками. Пищеварение полостное и пристеночное.
Задняя кишка имеет форму короткой трубки. Ее просвет выстлан кутикулой. У самцов задняя кишка открывается в так называемую клоаку, так как туда же открывается и отверстие полового аппарата. Из клоаки самца большей частью торчат наружу кутикулярные иголочки — спикулы — добавочные образования полового аппарата.
Кровеносная система отсутствует.
Дыхательной системы нет. Процесс дыхания носит анаэробный характер. Живя в кишечнике, нематоды получают энергию путем брожения накопленного гликогена.
Выделительная система протонефридиального характера, представлена отдельными экскреторными клетками. В боковых гиподермальных валиках проходят выделительные каналы, соединяющиеся в передней части тела и открывающиеся здесь наружу выделительной порой. Эти каналы проходят внутри единственной (иногда пары) Н-образной клетки, чья функция заключается главным образом в ионной регуляции.
Помимо этой части выделительной системы у нематод имеется и другая — фагоцитарная. По ходу каналов выделительной системы встречаются фагоцитарные клетки (“клетки накопления”), которые из полости тела захватывают бактерии, частицы отмерших клеток и т. п. Эти клетки крупные (две пары или более), располагаются чаще всего в передней трети тела нематод (их отростки сообщаются с выделительными каналами).
Половая система. Раздельнополые. Половые органы имени трубчатое строение. У самки парные, у самца — непарные.
Женские половые органы состоят из парных трубчатых яичников, трубчатого яйцевода и трубчатых маток, переходящих и непарную вагину и яйцемет. В полость матки проникают сперматозоиды после копуляции самки с самцом. Вокруг оплодотворенного яйца после оплодотворения формируется защитная оболочка. Она имеет сложную систему внеклеточных
мембран, обладающих энергетическим потенциалом, что обеспечивает дыхательную и защитную функцию для зародыша.
Мужские половые органы представлены непарным семенником, семяпроводом и семяизвергательным каналом, открывающимся в заднюю кишку.
Цикл развития. Самки и самцы аскариды паразитируют в тонких кишках человека, где и совершается оплодотворение. Самка откладывает здесь яйца, которые вместе с фекалиями выводятся наружу. Во внешней среде начинается дробление яиц, поскольку для этого необходимо наличие кислорода. Находясь в аэробных условиях, яйца аскариды развиваются в течение 21 суток до личиночной стадии. Такие яйца, содержащие уже оформившуюся “личинку”, становятся инвазионными, т. е. способными к дальнейшему развитию в теле хозяина (человека). Из этого видно, что яйца, только что выделившиеся наружу, не могут заразить хозяина. Их называют неинвазионными яйцами, т. е. неспособными вызвать паразитарное заболевание — инвазию.
При проглатывании инвазионных яиц личинки в кишечнике человека выходят из оболочек и активно проникают в кровеносные сосуды стенки кишки (рис. 3.49). С током крови по воротной вене они заносятся в печень, затем попадают в большой круг кровообращения и через нижнюю полую вену достигают правого предсердия; из предсердия в желудочек; из правого желудочка по легочной артерии личинки заносятся в капилляры, оплетающие легочные альвеолы, откуда током крови могут быть занесены в любой орган.
Рис. 3.49. Цикл развития человеческой аскариды

Из капилляров легких личинки “пробуравливаются” в альвеолы и по бронхам и трахее вместе с отделяемой от стенок дыхательных путей слизью попадают в ротовую полость. Личинки проглатываются со слюной и вторично оказываются в кишечнике, где и достигают половой зрелости.
Продолжительность миграции личинки от выхода из яйца до половозрелости не менее 75 дней.
Аскариды, не имея сильных органов прикрепления, тем не менее хорошо противостоят движениям кишечника, опираясь о его стенки своим изогнутым, пружинистым, напряженным телом. Взрослые аскариды выделяют ядовитые вещества и тем действуют на организм хозяина, вызывая расстройства кишечника, а также вредно влияют на нервную систему. Временами они обусловливают закупорку' кишечника (при массовых количествах), а в некоторых случаях прободение его стенки, результатом чего бывает перитонит (воспаление брюшины).
Личинки, если их много, могут быть причиной легочных заболеваний, так как они наносят многочисленные ранения стенкам альвеол.

Движение. Как много и как мало в данном слове. Для человека такое естественное свойство его опорно-двигательного аппарата открывает тысячи возможностей, и каждый наш день наполнен движением. И человек вовсе не заключен в своем теле, как в душной камере, напротив, люди вольны использовать ресурсы, данные им природой во всю мощь. Мышцы – удивительная ткань, возможности которой в условиях меняющихся нагрузок поражают воображение, а функциональные возможности скелетно-мышечной системы восхищают изящностью исполнения. А потому интересно было бы взглянуть на то, как же обеспечивается мышечная работа в целом.
В мышечной ткани происходит преобразование химической энергии в механическую работу. В качестве источника энергии химических связей используется АТФ, получаемая мышечными клетками в результате метаболических процессов.
Миоциты разных типов мышечной ткани обладают различным набором ферментов, а также отличаются по количеству митохондрий и миоглобина – белка, осуществляющего перенос кислорода. Мышечные волокна, выполняющие взрывную работу за короткое время покрывают необходимые энергетические затраты посредством анаэробного гликолиза, ферменты для осуществления которого присутствуют в клетках в норме в надлежащем количестве. За счет сравнительно низкого содержания миоглобина в клетках таких мышечных волокон, под микроскопом они выглядят светлыми и потому называются белыми волокнами. Им в противоположность существуют красные мышечные волокна, которые обеспечивают совершение продолжительной работы мышцами, и характеризуются более высоким содержанием миоглобина. Клетки красных мышечных волокон, которые, кроме прочего, составляют и сердечную мускулатуру, нуждаются в кислороде и потому имеют много митохондрий, а также богаты ферментами цикла Кребса и дыхательной цепи.
Глюконеогенез в мышечной ткани не протекает из-за отсутствия в ней нужных для его свершения ферментов. Продукты обмена веществ в мышечной ткани (лактат, возникающий при анаэробном гликолизе из пирувата, и аланин, являющийся продуктом трансаминирования из пирувата и аминокислот), пройдя циклы Кори и аланина, транспортируются по кровеносному руслу в печень для глюконеогенеза: в результате реакции повторного трансаминирования в печени возрастает количество необходимого пирувата. Гликоген печени при необходимости может стать источником глюкозы для мышечной ткани, что замыкает данный цикл. Синтез и разрушение гликогена в мышцах подчиняется гормональному контролю: инсулин способствует захвату глюкозы клетками и синтезу гликогена, тогда как катехоламины путем повышения уровня цАМФ стимулируют активность гликогенфосфорилазы. Катаболическое воздействие глюкокортикоидов ведет к разрушению мышечных белков и мобилизации аминокислот, которые в печени задействуются в глюконеогенезе.
Гидролитическое отщепление фосфатных групп от молекул АТФ дает мышечным клеткам необходимую для сокращения энергию. Еще одним макроэргическим фосфатом, используемым в мышцах, является креатинфосфат, отщепленная от которого креатинкиназой фосфатная группа переносится на АДФ. Неферментативно образующимся побочным продуктом превращения креатинфосфата является креатинин, который регулярно обнаруживается в крови и выводится через почки (суточное выводимое с мочой количество креатинина пропорционально мышечной массе). Таким образом, креатинфосфат представляет собой своего рода энергетический резерв, обеспечивая скорое восстановление количества молекул АТФ. У такой системы восстановления АТФ есть важное преимущество перед накоплением АТФ, заключающееся в том, что после превращения АТФ в АДФ вследствие разрыва фосфодиэфирной связи не создается невыгодного соотношения АТФ/АДФ, что могло бы негативно сказываться на функции АТФ из-за концентрационной зависимости от энергии Гиббса. Есть и еще одна система, служащая восстановлению АТФ в мышечной ткани, ключевым ее ферментом является аденилаткиназа (миокиназа), способная образовывать АТФ за счет АДФ, а также фосфорилировать АМФ до АДФ.
Строение мышечных волокон
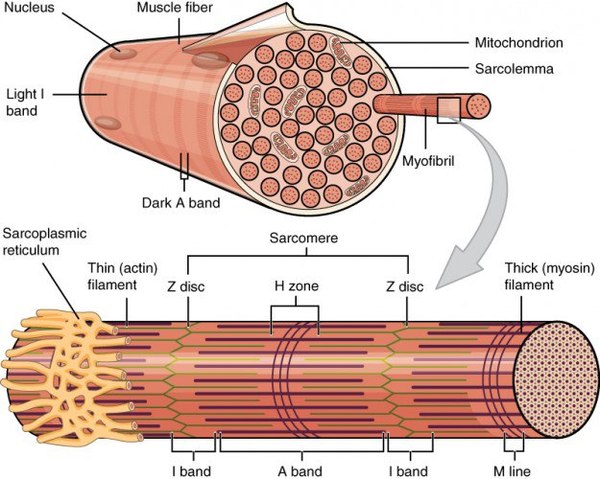
Клетки поперечно-полосатой мускулатуры отличаются от клеток гладкой мышечной ткани и ткани сердечной мышцы. Они образуют единый многоядерный синцитий. Клеточные ядра при этом смещены к краю клеток, а основное внутриклеточное пространство занято миофибриллами. Миофибриллы окружены саркоплазматическим ретикулумом, что достигается посредством формирования продольных и поперечных трубочек, а также лабиринтообразных впячиваний плазматической мембраны, благодаря чему возбуждение достигает этих участков. Плазматическая мембрана миоцитов – сарколемма – укреплена с внутренней стороны белками цитоскелета. В связывании с интегральными мембранными белками задействован белок дистрофин, мутации в гене которого приводят к развитию миодистрофии.
Скелетные мышцы структурно строго организованы в мышечные пучки, волокна, фибриллы и филаменты. Фибриллы поперечно-полосатых мышц состоят, прежде всего, из толстых миозиновых филаментов и тонких филаментов. Первый тип состоит из миозина и молекулярные моторные единицы. Второй тип филаментов включает F-актин и актин-связывающие белки – тропомиозин и тропонин. Головки тяжелых цепей миозина выдаются кнаружи и способны формировать связи с тонкими актиновыми филаментами. Актиновые филаменты, в свою очередь, закреплены на структурных белках, образующих так называемые Z-диски. Типичная поперечная исчерченность данного типа мышечной ткани, узнаваемая гистологически, создается благодаря устройству и расположению саркомера – функциональной единицы мышцы - который представляет собой участок миофибриллы между двумя Z-мембранами. Актиновые филаменты связываются как между собой, так и с Z-белками. Два других белка – титин и небулин – принимают участие в структурировании миофибрилл в процессе сокращения и расслабления. Титин прикрепляется к белкам Z-структуры и к М-линиям, сформированным миозином и структурными белками. Небулин закреплен в Z-структурах и исполняет регуляторную роль в построении тонких филаментов.
Несмотря на строгую организацию, скелетная мускулатура являет собой крайне гетерогенную систему относительно устройства и выполняемых функций. С одной стороны, это позволяет подстроиться мышцам под возлагаемую на них нагрузку путем разборки / увеличения количества саркомеров или миофибрилл, с другой стороны – обмен различными изоформами белков разных свойств и качеств обуславливает функциональную реорганизацию.
Мышечное сокращение
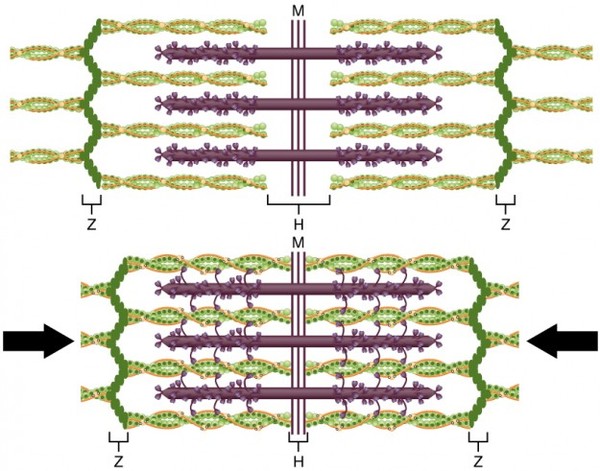
Схематичное строение саркомера.
Фиолетовым изображен миозин, головки его тяжелых цепей обращены к нитям актина и соединяются с ними. Движение головок миозина при сокращении ведет к подтягиванию актиновых филаментов к центру. Также обозначено прикрепление актиновых нитей к Z-дискам.
Сокращение мышц находится в зависимости от концентрации ионов кальция. Передача возбуждения на нейромышечные окончания поперечно-полосатых волокон ведет к деполяризации наружной мембраны и открытию кальциевых каналов в мембране саркоплазматического ретикулума, простирающегося через все саркомеры. Концентрация ионов кальция в цитозоле резко возрастает и они связываются с тропонином С. Это приводит к конформационным изменениям субъединиц тропонинового комплекса, что имеет следствием смещение позиции тропомиозина. Это событие делает возможным связывание участков цепей миозина с актином. Головки миозина расщепляют АТФ до АДФ и остатка фосфорной кислоты, а высвобождающаяся при этом энергия обеспечивает смену ее конформации. Свершившееся связывание ионов кальция с тропонином С является необходимым для взаимодействия миозина с актиновыми волокнами, которое влияет на изменение угла между легкой и тяжелой цепями миозина. Такой сдвиг оканчивается перемещением актинового филамента к центру саркомера. Скольжение тонких филаментов относительно толстых приводит к укорочению саркомеров и сокращению мышцы. После этого происходит экзергоническая смена конформации головки миозина, а продукты гидролиза АТФ выносятся из клетки. Для повторения цикла необходимо очередное внесение АТФ в систему. Если мотонейроны перестают получать раздражение извне, то в работу вступает АТФ-зависимый кальциевый насос, перекачивающий более не востребованные ионы кальция из цитоплазмы в цистерны саркоплазматического ретикулума, где они связываются с кальсеквестрином, обладающим высокой связывающей способностью. В связи с этим концентрация свободных ионов кальция снижается, что энергетически облегчает последующее поглощение этих ионов.
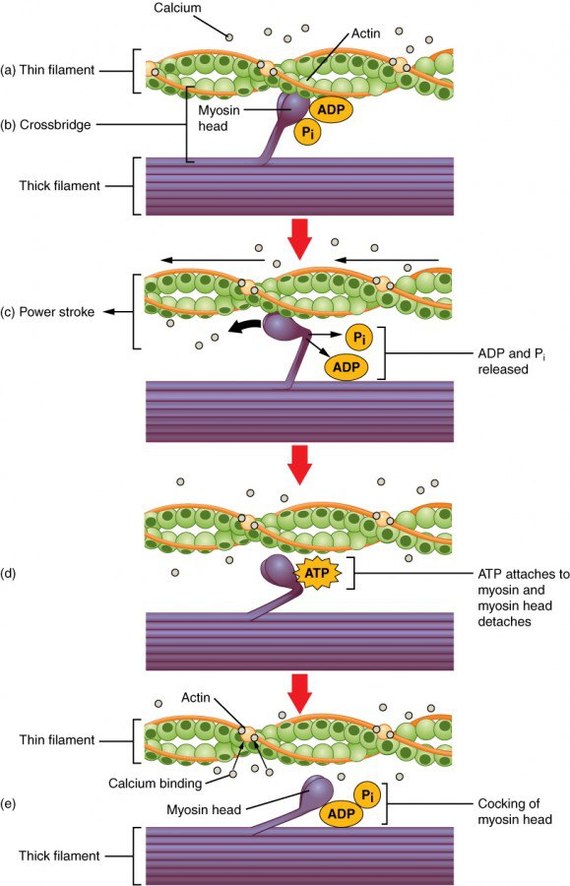
Схема мышечного сокращения.
Молекулярные компоненты системы, осуществляющей впоследствии высвобождение кальция из саркоплазматического ретикулума , известны только частично. Начало сигнальной цепочки опосредуют дигидропиридиновые рецепторы плазматической мембраны, которые сменяют свою конформацию под влиянием деполяризации мембраны. Это приводит к открытию кальциевых каналов и активации рианодиновых рецепторов терминальных цистерн саркоплазматического ретикулума. Преходящее повышение концентрации ионов кальция в цитоплазме мышечных клеток ведет и к метаболическим изменениям. Например, дефосфорилированная форма киназы гликогенфосфорилазы может активироваться комплексом кальмодулина с ионами кальция, а потому мышечное возбуждение связано с кратковременным разрушением гликогена.
На этом наше повествование не оканчивается. В последующих постах обязательно подробнее рассмотрим биохимические превращения в мышцах, а также обратимся к особенностям, происходящим с мышцами при физических нагрузках.
Löffler, Petrides Biochemie und Pathobiochemie, Springer, 2007
Волков, Несен Биохимия мышечной деятельности, 2000
Читайте также:
