
Ядра крестцового отдела спинного мозга симпатический
Центральная нервная система
Чувствительные нервные узлы
Они лежат по ходу задних корешков спинного мозга (спинно-мозговые узлы), либо черепномозговых нервов (че репномозговые узлы). Развиваются из нервного гребня.
Спинномозговой узел (спинальный ганглий). Имеет веретеновидную форму. Относится к паренхиматозным органам и, соответственно, состоит из стромы и па рен химы. Строма представлена соединительнотканной кап сулой, от которой вглубь узла отходят прослойки рыхлой волокнистой соединительной ткани с кровеносными сосудами. Паренхима узла представлена нервной тканью, состоя щей из нервных и глиальных клеток. Тела нервных клеток крупные, округлой формы, лежат группами на периферии узла. Это псевдоуниполярные (ложноодноотростчатые) рецепторные (чувствительные) нейроны. Поскольку аксон и дендрит, отходя от тела нейрона, плотно прилежат друг к другу, создаётся ложное впечатление, что отросток только один. За тем, на некотором расстоянии от тела нейрона, аксон и дендрит Т образно расходятся. При этом дендрит идёт в составе смешанных спинномозговых нервов на периферию и заканчивается там рецептором. Аксон входит в спинной мозг через задний корешок и несёт нервные импульсы к нейронам спинного либо головного мозга. Дендриты и аксоны нейронов спинномозгового узла окружены миелиновыми оболочками, а тело нейрона – слоем олигодендроцитов (мантийные глиоциты) с базальной мембраной, выполняющих трофическую и разграничительную функции. Снаружи слоя олигодендроцитов каждый нейрон спинномозгового узла окружён также и тонкой соединительнотканной оболочкой.
В ЦНС выделяют серое и белое вещество. Серое вещество образовано телами нейронов и окружающими их глиоцитами, а белое вещество – нервными волокнами. Выделяют два способа организации нейронов в ЦНС: ядерный, когда тела нейронов располагаются в виде скоплений, ядер (спинной мозг, ствол головного мозга) и экранный, когда тела нейронов располагаются послойно (в коре мозга и мозжечка). Ансамбли нейронов формируют нервные центры, соответственно, ядерного и экранного типа.
Спинной мозг располагается в позвоночном канале и имеет сегментарное строение. С каждым его сегментом связана пара передних (вентральных) и пара задних (дорзальных) корешков.
Это паренхиматозный орган, состоящий из стромы и паренхимы. Снаружи спинной мозг окружен тремя мозговыми оболочками (выполняют функции капсулы), от которых отходят прослойки рыхлой волокнистой соединительной ткани с кровеносными сосудами, питающими спинной мозг. Паренхима образована нервной тканью. Спинной мозг состоит из двух симметричных половин, разделённых спереди срединной щелью, а сзади – соединительнотканной перегородкой.
В сером веществе лежат тела нервных клеток спинного мозга. Они образуют скопления – ядра. Все нейроны спинного мозга мультиполярные. Среди них можно выделить следующие виды клеток: корешковые клетки, нейриты которых покидают спинной мозг в составе его передних корешков, пучковые клетки, аксоны которых выходят в белое вещество и идут в другие сегменты спинного мозга или головной мозг, и внутренние клетки, аксоны которых заканчиваются в пределах серого вещества своего сегмента спинного мозга.
В задних рогах расположены мелкие вставочные ней роны. Они воспринимают возбуждение от чувствительных нейронов спинальных ганглиев и передают его на эфферентные нейроны передних рогов спинного мозга, либо на другие вставочные нейроны спинного мозга, либо по восходящим проводящим путям на нейроны головного мозга. В зад них рогах выделяют несколько структурных образований: губчатый слой, желатинозное вещество, собственное ядро заднего рога, ядро Кларка. По медиаторной природе нейроны задних рогов пептидергические и ГАМКергические.
В грудном и поясничном отделах спинного мозга расположены боковые рога. Здесь различают медиальное и латеральное ядра, которые являются центрами симпатической нервной системы и образованы холинергическими нейронами. Их аксоны образуют синапсы на эфферентных аминергических нейронах, лежащих в симпатических узлах (паравертебральных, чревных и др.). В боковых рогах крестцового отдела спинного мозга находятся центральные нейроны (ядра) пара симпатической нервной системы.
В передних рогах расположены самые крупные ней роны спинного мозга (100-150 мкм). Это эффекторные нейроны (мотонейроны). Они передают возбуждение на скелетную мускулатуру и заканчиваются моторными бляшками на поперечнополосатых мышечных волокнах. Их называют корешковыми нейронами, поскольку их аксоны формируют передние корешки спинного мозга.
В настоящее время получило также распространение разделение серого вещества спинного мозга на 10 колонок (они хорошо видны на продольных срезах мозга), пластины по Рекседу. Они соответствуют топографии ядер серого вещества. I-V пластины находятся в задних рогах, VI-VII – в промежуточной зоне, VIII-IX – в передних рогах, а X пластина – вокруг центрального канала.
Белое вещество более светлое, расположено по периферии спинного мозга. Оно образовано преимущественно миелиновыми нервными волокнами, формирующими проводящие пути, связывающие различные сегменты спинного мозга, либо спинной мозг с головным. В белом веществе различают три пары канатиков или столбов: передних, задних и боковых. Они расположены, соответственно, между задними рогами, между задним и передним рогами и между передними рогами. Подробное описание ядер серого вещества и проводящих путей белого вещества спинного мозга даётся в соответствующем разделе анатомии человека.
Глиоциты спинного мозга. Эпендимоциты выстилают спинномозговой канал; отростки, отходящие от их базальной части, проходят через весь спинной мозг и входят в состав его наружной пограничной глиальной мембраны. Эпендимоциты могут выполнять также эндокринную функцию, выделяя биологи чески активные вещества в спинномозговую жидкость, циркулирующую по каналам и желудочкам мозга. Протоплазматические и волокнистые астроциты образуют глиальные мембраны внутри и на поверхности спинного мозга, вокруг кровеносных сосудов, питающих спинной мозг, выполняя опорную и разграничительную (изолирующую) функции. Олигодендроциты образуют оболочки вокруг крупных нейронов (сателлитные клетки) и отростков нервных клеток (леммоциты).
Вегетативная нервная система (ВНС)
Если соматическая нервная система иннервирует только скелетную мускулатуру, то вегетативная – все органы, включая и кровеносные сосуды скелетных мышц. Соответственно, она регулирует кровяное давление, микроциркуляцию, пищеварение, температуру тела, потоотделение, обмен веществ. ВНС ещё называют автономной, поскольку она, в отличие от соматической, не подчиняется сознанию, ею нельзя управлять усилием воли.
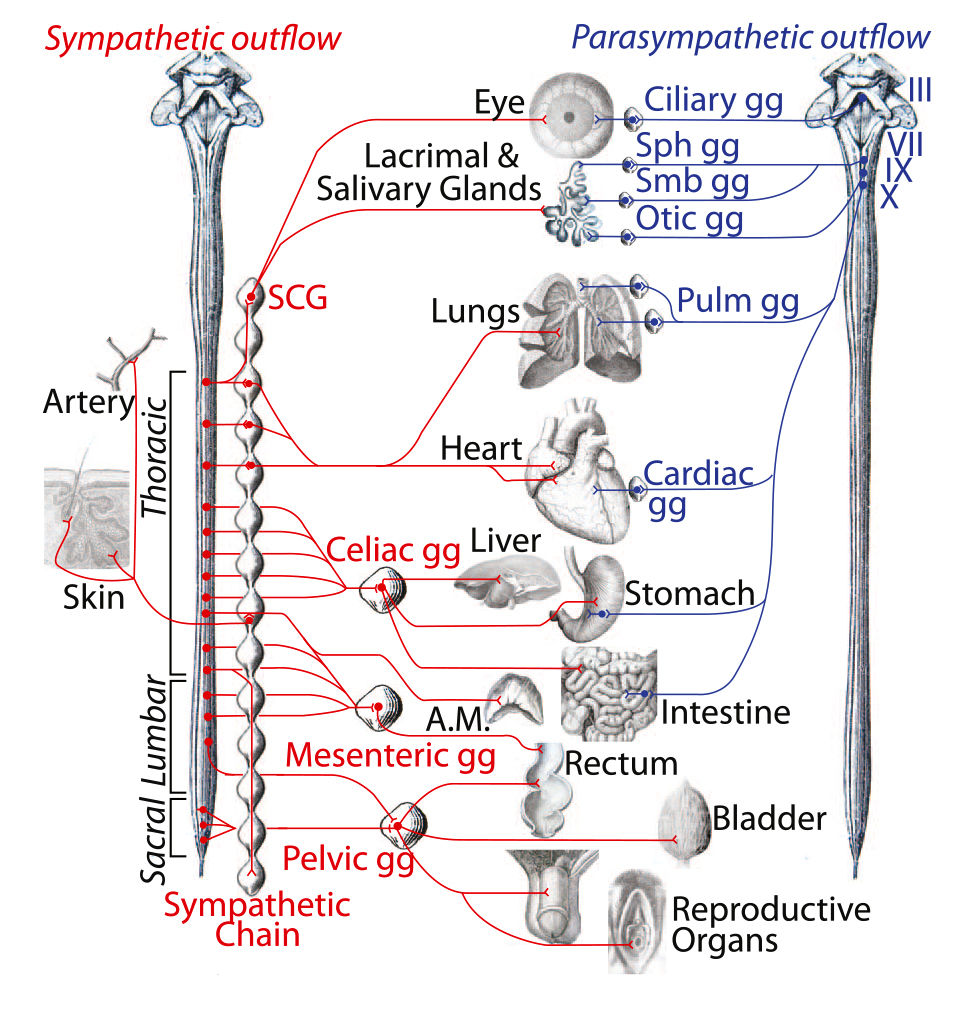
По своей функции и строению вегетативная нервная система делится на симпатическую, парасимпатическую и метасимпатическую. Анатомически вегетативная нервная система состоит из двух отделов: центрального и периферического. Центральный отдел представлен ядрами, лежащими в головном и спинном мозге, а периферический – нервными стволами, узлами (ганглиями), сплетениями и нервными окончаниями.
Центральный отдел симпатическойнервной системы расположен в боковых рогах грудного и поясничного отделов спинного мозга, а парасимпатической – в боковых рогах крестцового отдела спинного мозга, а также в среднем и продолговатом отделах ствола головного мозга. Аксоны рас положенных в центральном отделе нейронов образуют преганглионарные нервные волокна. Они покрыты миелиновой оболочкой и заканчиваются синапсами на нейронах вегетативных нервных ганглиев. Последние лежат как вне органов (экстрамуральные), так и в стенке внутренних органов (интрамуральные). Ганглии окружены соединительнотканной капсулой, от которой отходят прослойки рыхлой волокнистой соединительной ткани, окружающей каждый нейрон узла.
Ганглии симпатической нервной системылежат либо двумя цепочками вдоль спинного мозга – паравертебральные ганглии, либо в брюшной полости – чревный и брыжеечныеганглии. Медиатором в их нейронах является норадреналин. Кроме нейронов трёх типов Догеля, здесь обнаружены мелкие интенсивно флюоресцирующие клетки (МИФ-клетки). Предполагают, что они выполняют тормозную функцию.
Ганглии парасимпатической нервной системы лежат обычно на поверхности или внутри органов. Они также содержат клетки трёх типов Догеля, медиатором которых является преимущественно ацетилхолин.
Каждый орган обычно иннервируется двумя отделами ВНС: симпатическим и парасимпатическим, которые оказывают на него противоположное воздействие. Например, симпатическая нервная система активируется при остром стрессе и ускоряет сердцебиение, суживает кровеносные сосуды и повышает кровяное давление, но тормозит пищеварение, а парасимпатическая нервная система замедляет сердцебиение, расширяет кровеносные сосуды и снижает кровяное давление, но стимулирует пищеварение. Последняя активируется при отдыхе и восстановлении ресурсов организма.
Рефлекторная дуга вегетативной нервной системы. В отличие от соматической, эфферентная часть рефлекторной дуги вегетативной нервной системы всегда двухнейронная: первый нейрон лежит в ЦНС, а второй – в вегетативном ганглии. Аксоны центральных вегетативных нейронов образуют преганглионарные волокна и заканчиваются синап сами на эфферентных нейронах, которые находятся в вегетативных ганглиях. Нейроны последних образуют постганглионарные волокна, которые заканчиваются эфферентными нервными окончаниями на клетках рабочих органов. Афферентные (чувствительные) нейроны лежат, в спинальных ганглиях, либо в черепномозговых узлах.
Метасимпатическая нервная система.В её состав входят нейроны трёх типов Догеля, тела которых расположены в интрамуральных ганглиях. Нейроны 1-го типа – длинноаксонные,имеют короткие дендриты и длинный аксон, уходящий за пределы ганглия. Это эфферентные нейроны, их аксоны образуют безмиелиновые постганглионарные нервные волокна и заканчиваются на рабочих клетках (гладкомышечных и секреторных). Нейроны 2-го типа – равноотростчатые,афферентные; их аксоны заканчиваются на нейронах 1-го типа. Между ними находятся нейроны 3-го типа – вставочные, ассоциативные, они посылают свои отростки клеткам I типа и в другие ганглии.
Метасимпатическая нервная система регулируется как симпатической, так и парасимпатической нервной системой, но не относится к ним и обладает относительной независимостью. Она образует местные рефлекторные дуги, и может осуществлять нервную регуляцию даже изолированного органа. Этим обеспечивается относительная автономность деятельности внутренних органов.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Вегетативная (автономная, висцеральная) нервная система – эта неотъемлемая часть нервной системы человека. Ее основной функцией является обеспечение деятельности внутренних органов. Она состоит из двух отделов, симпатического и парасимпатического, которые обеспечивают противоположные влияния на органы человека. Работа вегетативной нервной системы очень сложна и относительно автономна, почти не подчиняется воле человека. Давайте познакомимся поближе со строением и функциями симпатического и парасимпатического отделов вегетативной нервной системы.
Понятие о вегетативной нервной системе

Автономная нервная система состоит из нервных клеток и их отростков. Как и у обычной нервной системы человека, у вегетативной выделяют два отдела:
- центральный;
- периферический.
Центральная часть осуществляет контроль над функциями внутренних органов, это руководящий отдел. В нем нет четкого деления на противоположные по сфере влияния части. Он задействован в работе всегда, круглые сутки.
Периферическая часть вегетативной нервной системы представлена симпатическим и парасимпатическим отделами. Структуры последних есть практически в каждом внутреннем органе. Работают отделы одновременно, но, в зависимости от того, что требуется в данный момент от организма, какой-то один оказывается превалирующим. Именно разнонаправленные влияния симпатического и парасимпатического отделов позволяют организму человека приспосабливаться к постоянно меняющимся условиям окружающей среды.
Функции вегетативной нервной системы:
- поддержание постоянства внутренней среды (гомеостаза);
- обеспечение всей физической и психической деятельности организма.
Вам предстоит физическая нагрузка? С помощью вегетативной нервной системы артериальное давление и сердечная деятельность обеспечат достаточный минутный объем кровообращения. У Вас отдых, и частые сердечные сокращения совсем ни к чему? Висцеральная (вегетативная) нервная система заставит сердце сокращаться медленнее.
Центральный отдел
Эта часть автономной нервной системы представляет собой различные структуры головного мозга. Она, получается, как бы разбросана по всему мозгу. В центральном отделе выделяют сегментарные и надсегментарные структуры. Все образования, относящиеся к надсегментарному отделу, объединяют под названием гипоталамо-лимбико-ретикулярный комплекс.

Гипоталамус – это структура головного мозга, расположенная в нижней его части, у основания. Нельзя сказать, что это область с четкими анатомическими границами. Гипоталамус плавно переходит в мозговую ткань других отделов мозга.
В целом гипоталамус состоит из скопления групп нервных клеток, ядер. Всего изучено 32 пары ядер. В гипоталамусе формируются нервные импульсы, которые по различным проводящим путям достигают других структур головного мозга. Эти импульсы управляют кровообращением, дыханием, пищеварением. В гипоталамусе расположены центры регуляции водно-солевого обмена, температуры тела, потоотделения, голода и насыщения, эмоций, полового влечения.
Кроме нервных импульсов, в гипоталамусе образуются вещества гормоноподобной структуры: рилизинг-факторы. С помощью этих веществ осуществляется регуляция деятельности молочных желез (лактация), надпочечников, половых желез, матки, щитовидной железы, роста, расщепления жиров, степени окраски кожи (пигментация). Все это возможно благодаря тесной связи гипоталамуса с гипофизом – главным эндокринным органом организма человека.
Таким образом, гипоталамус функционально связан со всеми отделами нервной и эндокринной систем.
Условно в гипоталамусе выделяют две зоны: трофотропную и эрготропную. Деятельность трофотропной зоны направлена на поддержание постоянства внутренней среды. Она связана с периодом отдыха, поддерживает процессы синтеза и утилизацию продуктов метаболизма. Свои основные влияния реализует через парасимпатический отдел вегетативной нервной системы. Стимуляция этой зоны гипоталамуса сопровождается усиленным потоотделением, слюноотделением, замедлением сердечных сокращений, снижением артериального давления, расширением сосудов, повышением перистальтики кишечника. Трофотропная зона расположена в передних отделах гипоталамуса. Эрготропная зона ответственна за приспособляемость организма к меняющимся условиям, обеспечивает адаптацию и реализуется через симпатический отдел вегетативной нервной системы. При этом повышается артериальное давление, ускоряется сердцебиение и дыхание, расширяются зрачки, повышается содержание сахара в крови, снижается перистальтика кишечника, тормозится мочеотделение и дефекация. Эрготропная зона занимает задние отделы гипоталамуса.
К этой структуре относят часть коры височной доли, гиппокамп, миндалину, обонятельную луковицу, обонятельный тракт, обонятельный бугорок, ретикулярную формацию, поясную извилину, свод, сосочковые тела. Лимбическая система участвует в формировании эмоций, памяти, мышления, обеспечивает пищевое и сексуальное поведение, регулирует цикл сна и бодрствования.
Для реализации всех этих влияний необходимо участие многих нервных клеток. Система функционирования очень сложна. Чтобы сформировалась определенная модель поведения человека, нужна интеграция многих ощущений с периферии, передача возбуждения одновременно к различным структурам головного мозга, как бы циркуляция нервных импульсов. Например, для того, чтобы ребенок запомнил названия времен года, необходима многократная активация таких структур, как гиппокамп, свод, сосочковые тела.
Эту часть вегетативной нервной системы называют сетчатой, потому что она, как сеть, оплетает все структуры головного мозга. Такое диффузное расположение позволяет ей участвовать в регуляции всех процессов в организме. Ретикулярная формация поддерживает кору головного мозга в тонусе, в постоянной готовности. Этим обеспечивается мгновенное включение нужных областей коры головного мозга. Особенно это важно для процессов восприятия, памяти, внимания и обучения.
Отдельные структуры ретикулярной формации ответственны за конкретные функции в организме. Например, есть дыхательный центр, который располагается в продолговатом мозге. Если он поражается по какой-либо причине, то самостоятельное дыхание становится невозможным. По аналогии есть центры сердечной деятельности, глотания, рвоты, кашля и так далее. Функционирование ретикулярной формации также основано на наличии многочисленных связей между нервными клетками.
В целом, все структуры центрального отдела вегетативной нервной системы взаимосвязаны посредством многонейронных связей. Только их согласованная деятельность позволяет реализовать жизненно важные функции вегетативной нервной системы.
Эта часть центрального отдела висцеральной нервной системы имеет четкое деление на симпатические и парасимпатические структуры. Симпатические структуры располагаются в грудо-поясничном отделе спинного мозга, а парасимпатические – в головном мозге и крестцовом отделе спинного мозга.
Симпатические центры локализуются в боковых рогах в следующих сегментах спинного мозга: С8, все грудные (12), L1, L2. Нейроны этой области участвуют в иннервации гладких мышц внутренних органов, внутренних мышц глаза (регуляция величины зрачка), желез (слезных, слюнных, потовых, бронхиальных, пищеварительных), кровеносных и лимфатических сосудов.
Содержит следующие образования в головном мозге:
- добавочное ядро глазодвигательного нерва (ядро Якубовича и Перлиа): контроль величины зрачка;
- слезное ядро: соответственно, регулирует слезоотделение;
- верхнее и нижнее слюноотделительные ядра: обеспечивают продукцию слюны;
- дорсальное ядро блуждающего нерва: обеспечивает парасимпатические влияния на внутренние органы (бронхи, сердце, желудок, кишечник, печень, поджелудочную железу).
Крестцовый отдел представлен нейронами боковых рогов сегментов S2-S4: они регулируют мочеиспускание и дефекацию, кровенаполнение сосудов половых органов.
Периферический отдел
Представлен симпатическим стволом, расположенным по обе стороны от позвоночника. Симпатический ствол – это два ряда (правый и левый) нервных узлов. Узлы имеют связь друг с другом в виде мостиков, перебрасывающихся между частями одной стороны и другой. То есть, ствол выглядит как цепочка из нервных комочков. В конце позвоночника два симпатических ствола соединяются в один непарный копчиковый узел. Всего различают 4 отдела симпатического ствола: шейный (3 узла), грудной (9-12 узлов), поясничный (2-7 узлов), крестцовый (4 узла и плюс один копчиковый).
В области симпатического ствола располагаются тела нейронов. К этим нейронам подходят волокна от нервных клеток боковых рогов симпатической части центрального отдела вегетативной нервной системы. Импульс может переключаться на нейронах симпатического ствола, а может проходить транзитом и переключаться на промежуточных узлах нервных клеток, расположенных или вдоль позвоночника или вдоль аорты. В дальнейшем волокна нервных клеток после переключения в узлах формируют плетения. В области шеи это сплетение вокруг сонных артерий, в грудной полости это сердечное и легочное сплетения, в брюшной – солнечное (чревное), верхнее брыжеечное, нижнее брыжеечное, брюшное аортальное, верхнее и нижнее подчревные. Эти крупные сплетения делятся на более мелкие, от которых вегетативные волокна движутся к иннервируемым органам.
От центральных парасимпатических центров, расположенных в головном мозге, импульсы идут в составе черепно-мозговых нервов (соответственно глазодвигательного, лицевого и тройничного, языкоглоточного и блуждающего). Поскольку блуждающий нерв участвует в иннервации внутренних органов, то в его составе волокна достигают глотки, гортани, пищевода, желудка, трахеи, бронхов, сердца, печени, поджелудочной железы, кишечника. Выходит, что большинство внутренних органов получает парасимпатические импульсы из системы разветвлений всего лишь одного нерва: блуждающего.
От крестцовых отделов парасимпатической части центральной висцеральной нервной системы нервные волокна идут в составе тазовых внутренностных нервов, достигают органов малого таза (мочевого пузыря, мочеиспускательного канала, прямой кишки, семенных пузырьков, предстательной железы, матки, влагалища, части кишечника). В стенках органов импульс переключается в нервных узлах, и короткие нервные веточки непосредственно контактируют с иннервируемой областью.
Выделяется как некий отдельно существующий отдел вегетативной нервной системы. Выявляется преимущественно в стенках внутренних органов, обладающих способностью к сокращению (сердце, кишечник, мочеточник и другие). Состоит из микроузлов и волокон, образующих нервное сплетение в толще органа. Структуры метасимпатической вегетативной нервной системы могут реагировать как на симпатические, так и на парасимпатические влияния. Но, кроме того, доказана и их способность работать автономно. Считается, что перистальтическая волна в кишечнике – это результат функционирования метасимпатической вегетативной нервной системы, а симпатический и парасимпатический отделы лишь регулируют силу перистальтики.
Как работают симпатический и парасимпатический отделы?

Закон противоположностей
Симпатическая иннервация обеспечивает:
Парасимпатическая иннервация действует следующим образом:
Из общей закономерности есть исключения. В организме человека есть структуры, которые имеют только симпатическую иннервацию. Это стенки сосудов, потовые железы и мозговой слой надпочечников. На них парасимпатические влияния не распространяются.
Обычно в организме здорового человека влияния обоих отделов находятся в состоянии оптимального равновесия. Возможно незначительное преобладание одного из них, что тоже является вариантом нормы. Функциональное преобладание возбудимости симпатического отдела называется симпатикотонией, а парасимпатического отдела – ваготонией. Некоторые возрастные периоды человека сопровождаются повышением или снижением активности обоих отделов (например, в подростковый период повышается активность, а в период старости снижается). Если наблюдается превалирующая роль симпатического отдела, то это проявляется блеском в глазах, широкими зрачками, склонностью к повышенному артериальному давлению, запорам, избыточной тревожностью и инициативностью. Ваготоническое действие проявляется узкими зрачками, склонностью к пониженному артериальному давлению и обморокам, нерешительностью, избыточной массой тела.
Таким образом, из выше изложенного становится понятно, что автономная нервная система со своими противоположно направленными отделами обеспечивает жизнедеятельность человека. Причем работают все структуры согласованно и скоординировано. Деятельность симпатического и парасимпатического отделов не контролируется человеческим мышлением. Это именно тот случай, когда природа оказалась умнее человека. У нас есть возможность заниматься профессиональной деятельностью, мыслить, творить, оставлять себе время на небольшие слабости, будучи уверенным, что собственный организм не подведет. Внутренние органы будут трудиться даже тогда, когда мы отдыхаем. И это все благодаря вегетативной нервной системе.

В течение столетия считалось, что черепные и крестцовые висцеральные нервы позвоночника — родственные образования. Согласно этому, крестцовые преганглионарные нейроны, как и ганглии, расположенные в области малого таза и контролирующие функции прямой кишки, мочевого пузыря, половой системы, считаются парасимпатическими. В этой публикации раскрываются 15 фенотипических и онтогенетических особенностей, которые отличают пре- и постганглионарные нейроны краниального отдела парасимпатической системы от симпатических нейронов грудопоясничного отдела у мышей. Каждая из особенностей доказывает, что крестцовый отдел неотличим от грудопоясничного. Таким образом, парасимпатическая нервная система получает импульсы исключительно от черепных нервов, а симпатическая нервная система от спинномозговых, в том числе от торакальных и крестцовых. Это упрощенное, двухкомпонентное строение, предлагает новую концепцию нейрофизиологии тазового дна, а также эволюции и развития вегетативной нервной системы.
Отнесение крестцового отдела к парасимпатической вегетативной нервной системе — как и связь черепного отдела с крестцовым — имеет древнюю, хотя и простую историю: это убеждение, уходящее корнями в работу Уолтера Гаскелла [1], было документально оформлено Лэнгли [2], и с тех пор стало общепринятым [3]. Аргументом послужили некоторые сходства крестцового отдела с краниальным: (I) анатомическое — область иннервации, в отличие от грудопоясничного отдела, менее рассеяна, отличается от него отсутствием иннервации конечностей, а также недостатком проекций на паравертебральный симпатический ствол [1] (II); физиологическое — влияние на некоторые органы противоположно грудопоясничному отделу [4]; и (III), фармакологическое — повсеместная чувствительность к антагонистам мускариновых рецепторов [2]. Однако, не хватало анализа клеточного фенотипа. В этой статье исследователи определили дифференциальные генетические признаки и взаимосвязи парасимпатических и симпатических нейронов, как пре-, так и постганглионарных. Когда они рассмотрели крестцовый отдел вегетативной нервной системы мышей с такого ракурса, то обнаружили, что его лучше отнести к симпатической, а не к парасимпатической системе.
Черепные парасимпатические преганглионарные нейроны образуются в прогениторной области заднего мозга “pMNv”[5], экспрессирующей гомеоген Phox2band и, ко всему прочему, формирующей бранхиомоторные нейроны [6]. Постмитотические предшественники мигрируют дорсально [7] с образованием ядер (таких как дорсальное двигательное ядро блуждающего нерва) и через дорсолатеральные точки выхода выступают в качестве ветвей некоторых черепных нервов, иннервирующих парасимпатические и энтеральные ганглии. В противоположность этому, симпатические преганглионарные нейроны грудного и верхнего поясничного отдела (далее просто “грудного”), предположительно, имеют общее происхождение с соматическими мотонейронами [8, 9]. Подразумевается, что они образуются в прогениторной области pMN (дорсальнее р3) — то есть из прогениторных клеток, которые экспрессируют основной фактор транскрипции OLIG2 типа спираль-петля-спираль (bHLH) [10]. Симпатические преганглионарные предшественники затем отделяются от соматических мотонейронов, формируя интермедиолатеральный столб у млекопитающих [11], затем вступают в вентральные корешки спинномозговых нервов вместе с аксонами соматических мотонейронов, и с помощью белых соединительных ветвей образуют синапсы на нейронах паравертебральных и превертебральных симпатических ганглиев.
Исследователи стремились сравнить генетический состав и взаимосвязи нижних поясничных и крестцовых преганглионарных нейронов (ниже просто “крестцовых”) с черепными (парасимпатическими) и грудными (симпатическими). В качестве представителя черепных преганглионарных нейронов они избрали дорсальное двигательное ядро блуждающего нерва, скопление нейронов, заметное уже на 13–14 день эмбрионального развития (E13.5) и экспрессирующее везикулярный переносчик ацетилхолина (VAChT) (рис. 1В). Грудные и крестцовые преганглионарные нейроны, которые образуют медиолатеральный столб спинного мозга, несмотря на их возможную холинергическую природу на данном этапе не экспрессируют VAChT. Для того, чтобы определить их локализацию, был использован общий маркер — синтаза оксида азота (NOS) [12] (рис.1, A и B), который отсутствовал в дорсальном двигательном ядре блуждающего нерва на E13.5 (рис.1В) или позже. Таким образом, экспрессия NOS характерна для грудных и крестцовых, но не для черепных преганглионарных нейронов.
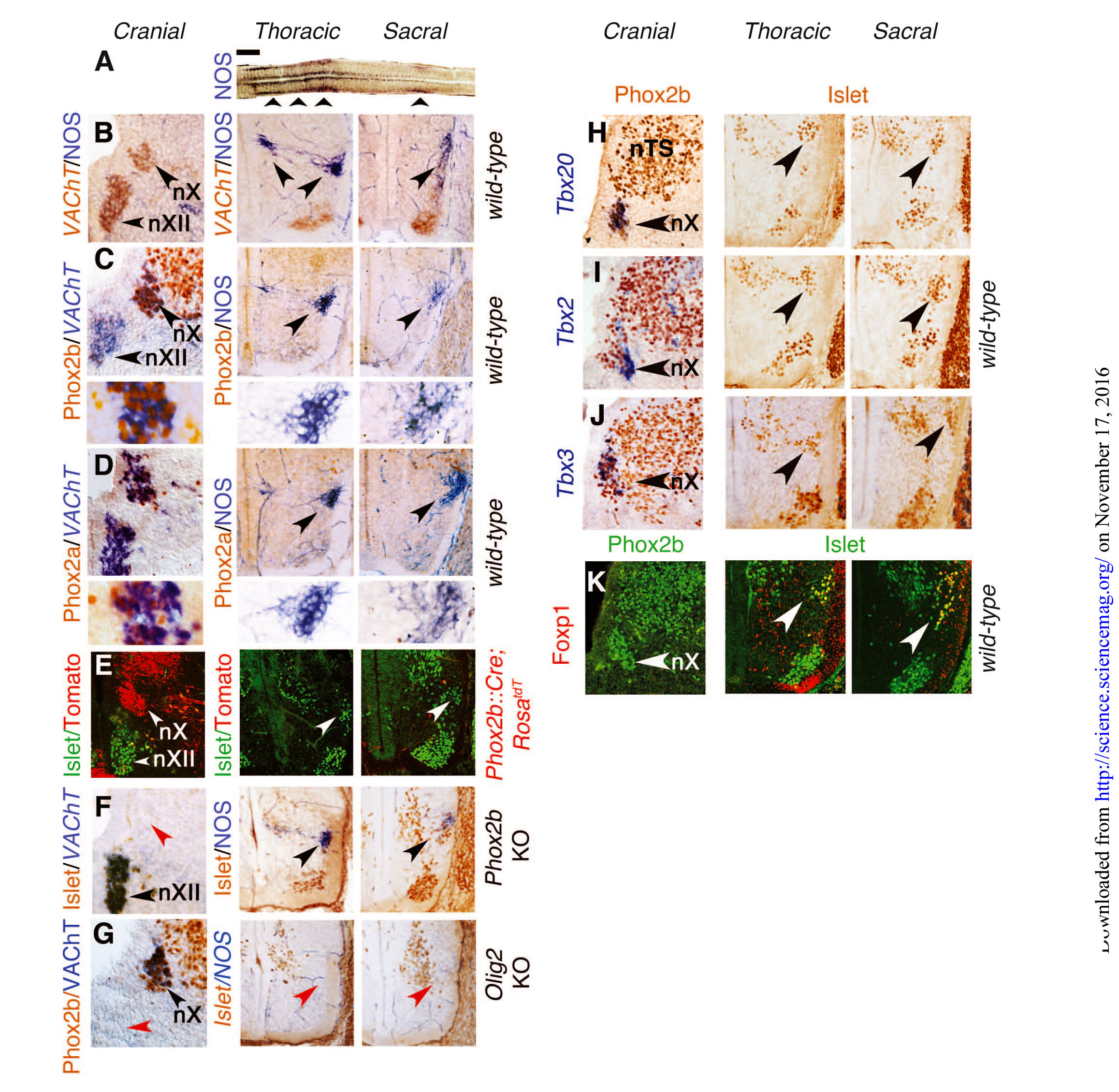
В отличие от черепных (парасимпатических) преганглионарных нейронов, грудные (симпатические) преганглионарные нейроны не только не экспрессируют Phox2b или его паралог Phox2a на E13.5, но также образуются из Phox2b-негативных прогениторов, а вместо Phox2b их дифференцировка (рис. 1, от C до F, левый и средний столбцы) зависит от OLIG2 (рис. 1G). Крестцовые преганглионарные нейроны в этом плане схожи с грудными (рис. 1, от С до G, средний и правый столбцы). На E13.5, экспрессия факторов транскрипции Tbx20, Tbx2 и Tbx3 семейства T-box была выражена в черепных (парасимпатических) нейронах, но отсутствовала как в грудных (симпатических), так и в крестцовых преганглионарных нейронах (рис. 1, от Н до J). Фактор транскрипции Foxp1 семейства F-box — определяющий фактор преганглионарных нейронов грудного отдела [13] — экспрессировали крестцовые, но не черепные преганглионарные нейроны (рис. 1К). Различия в экспрессии Phox2b, Tbx20 и FoxP1 между черепными и всеми спинномозговыми преганглионарными нейронами грудного и крестцового отделов, также наблюдается на E16.5. В целом, онтогенез и транскрипционные характеристики крестцовых преганглионарных нейронов были неотличимы от грудных, а, следовательно, и от симпатических нейронов.
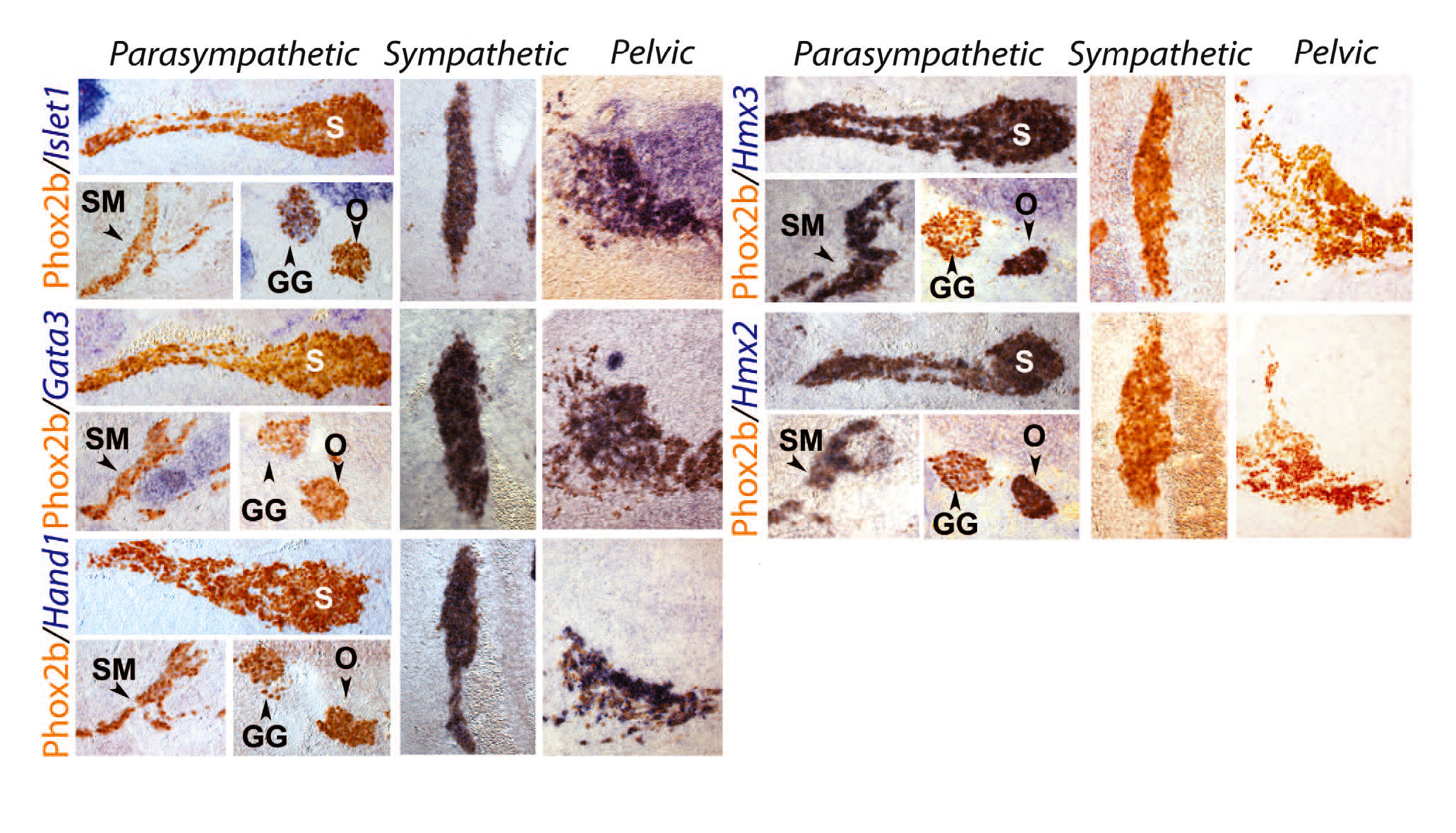
Грудные и крестцовые преганглионарные нейроны располагаются в медиолатеральной области спинного мозга, их аксоны выходят вентрально, в то время как топография черепных преганглионарных нейронов менее систематизирована, а их аксоны выходят дорсально. Эти сходства крестцового отдела с грудным, а также их отличия от краниального противоречат представлению о краниосакральном сходстве с момента его первого описания [1].
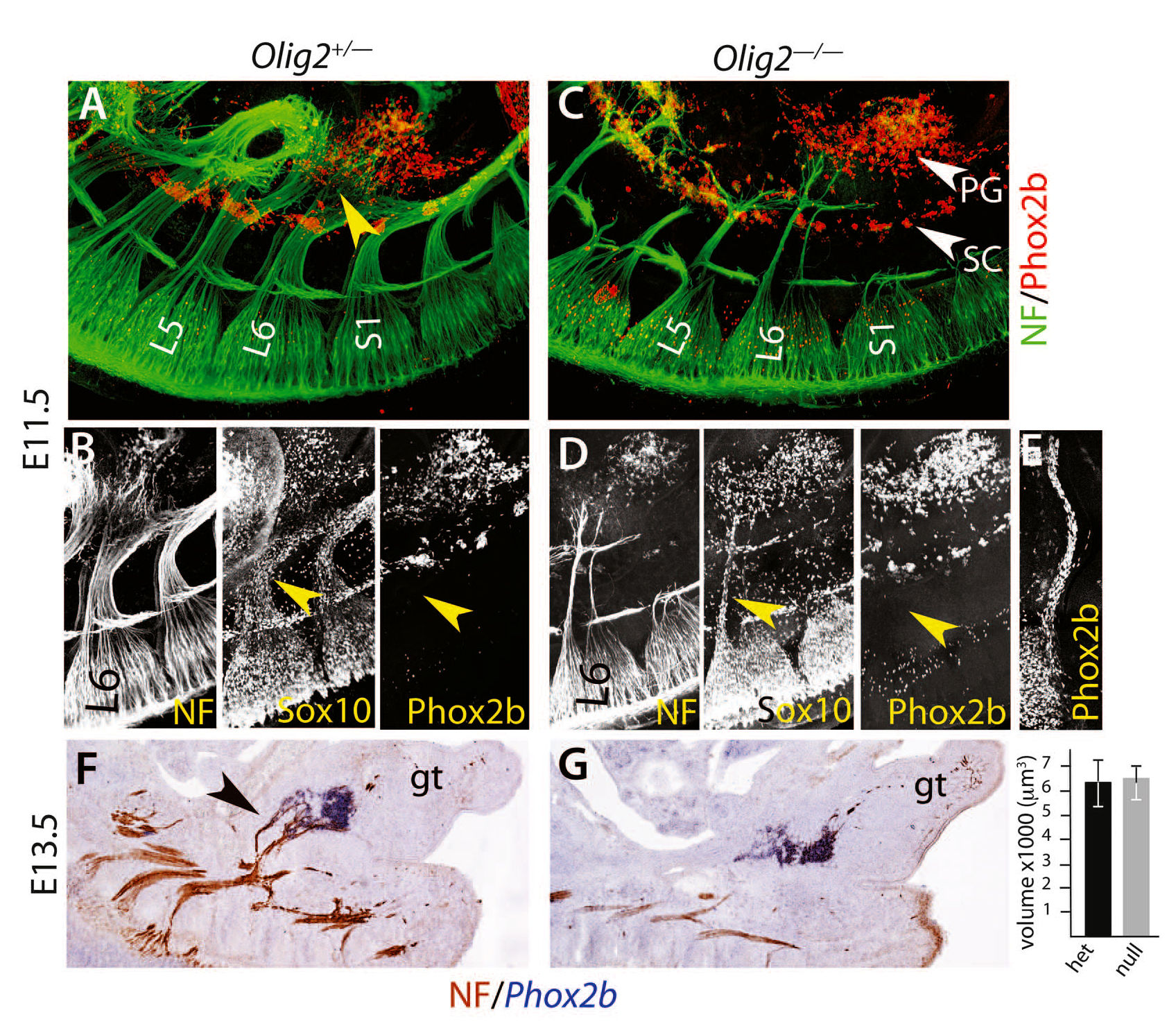
Наконец, опираясь на различия в режимах развития симпатических и парасимпатических ганглиев был проверен тазовый ганглий. Парасимпатические ганглии, в отличии от симпатических, возникают в результате миграции предшественников шванновских клеток Sox10+/Phox2b+ вдоль их будущего преганглионарного нерва по направлению к месту формирования ганглия и не образуют его, если эти нервы отсутствуют [24, 25]. На E11.5 пояснично-крестцовое сплетение, которое дает начало тазовому нерву, распростерло волокна, которые достигли бокового и рострального краев зачатка тазового ганглия и большинство из которых уже расположилось далеко впереди него (рис. 3А и видео S1). Эти волокна были покрыты клетками, экспрессирующими Sox10+, но (рис. 3B), в отличие от черепно-мозговых нервов, которые образуют парасимпатические ганглии на той же стадии, но не экспрессируют Phox2b (рис. 3E). Удаление всех моторных волокон у OLIG2-/- эмбрионов оставило только два тонких, предположительно, сенсорных, выступа от крестцового сплетения (рис. 3C), а также сильно уменьшило количество клеток с Phox2b+ (рис. 3D и рис. S10). Несмотря на эту массовую атрофию, тазовый ганглий оказался интактен (рис. 3C видео S2). Это было количественно проверено на E13.5 (рис. 3, F и G).
Хотя на 50% это клетки постганглионарного тазового нерва, тазовый ганглий формируется раньше и независимо от нерва, как и подобает симпатическому ганглию, а не парасимпатическому. Таким образом, крестцовый отдел вегетативной нервной системы является нижним форпостом симпатического отдела (рис. 4), а сама вегетативная нервная система разделена на краниальную и спинальную части, в соответствии с определенными эволюционными теориями [26]. Это новое понимание анатомии учитывает многие данные, которые противоречили предыдущим. Например, хотя обычно схемы представляют крестцовый путь к прямой кишке как дисинаптический, т.е. вагус-подобный [3], на самом деле он преимущественно [27, 28] трисинаптический, т.е. симпато-подобный [29]. Несмотря на догму об антагонизме между поясничным и крестцовым отделами, проявляющемся действием на мышцу-детрузор мочевого пузыря, в эксперименте торможение поясничного отдела отсутствует [4] или имеет сомнительную функциональную значимость [30]. Синергизм поясничного и крестцового отделов, проявляющийся расширением сосудов во внешних половых органах [29] показывает преемственность действия, а не антагонизм, как предлагает старая модель нервных импульсов грудопоясничного и крестцового отделов.
Симпатическая идентичность всех крестцовых и тазовых вегетативных нейронов, которую раскрывают новые данные, представляет собой основу для открытий в области нейроанатомии и физиологии таза.
- W. H. Gaskell, J. Physiol. 7, 1–80.9 (1886).
- J. N. Langley, The Autonomic Nervous System: Part I (W. Heffer, Cambridge, 1921).
- E. Kandel, J. Schwartz, T. Jessell, S. Siegelbaum, A. J. Hudspeth, Principles of Neural Science, Fifth Edition (McGraw-Hill Professional, 2012).
- J. N. Langley, H. K. Anderson, J. Physiol. 19, 71–139 (1895). 5. J. Briscoe et al., Nature 398, 622–627 (1999).
- A. Pattyn, M. Hirsch, C. Goridis, J. F. Brunet, Development 127, 1349–1358 (2000).
- S. Guthrie, Nat. Rev. Neurosci. 8, 859–871 (2007).
- Prasad, M. Hollyday, J. Comp. Neurol. 307, 237–258 (1991).
- A. Markham, J. E. Vaughn, J. Neurobiol. 22, 811–822 (1991).
- W. A. Alaynick, T. M. Jessell, S. L. Pfaff, Cell 146, 178–178.e1 (2011).
- P. E. Phelps, R. P. Barber, J. E. Vaughn, J. Comp. Neurol. 330, 1–14 (1993).
- C. R. Anderson, Neurosci. Lett. 139, 280–284 (1992).
- J. S. Dasen, A. De Camilli, B. Wang, P. W. Tucker, T. M. Jessell, Cell 134, 304–316 (2008).
- J. R. Keast, Int. Rev. Cytol. 248, 141–208 (2006).
- J. R. Keast, Neuroscience 66, 655–662 (1995).
- Kuntz, R. L. Moseley, J. Comp. Neurol. 64, 63–75 (1936).
- W. C. De Groat, A. M. Booth, J. Krier, in Integrative Functions of the Autonomic Nervous System, C. M. Brooks, K. Koizumi, A. Sato, Eds. (University of Tokyo Press, Tokyo, 1979), pp. 234–245.
- U. Ernsberger, H. Rohrer, Cell Tissue Res. 297, 339–361 (1999).
- K. Huber et al., Dev. Biol. 380, 286–298 (2013).
- K. Tsarovina et al., Development 131, 4775–4786 (2004).
- E. Doxakis, L. Howard, H. Rohrer, A. M. Davies, EMBO Rep. 9, 1041–1047 (2008).
- L. Huber, M. Ferdin, J. Holzmann, J. Stubbusch, H. Rohrer, Dev. Biol. 363, 219–233 (2012).
- C. L. Yntema, W. S. Hammond, J. Exp. Zool. 129, 375–413 (1955).
- V. Dyachuk et al., Science 345, 82–87 (2014).
- Espinosa-Medina et al., Science 345, 87–90 (2014).
- S. Nilsson, in Autonomic Nerve Function in the Vertebrates, Zoophysiology, vol. 13, D. S. Farner, Ed. (Springer-Verlag, New York, 1983), chap. 2.
- C. Olsson et al., J. Comp. Neurol. 496, 787–801 (2006). 28. K. Fukai, H. Fukuda, J. Physiol. 362, 69–78 (1985).
- W. Jänig, The Integrative Action of the Autonomic Nervous System: Neurobiology of Homeostatis (Cambridge Univ. Press, Cambridge, UK, 2006).
- W. C. de Groat, W. R. Saum, J. Physiol. 220, 297–314 (1972).
- A. Pattyn, X. Morin, H. Cremer, C. Goridis, J. F. Brunet, Nature 399, 366–370 (1999).
Читайте также:
