
Кортико спинальный тракт это
Двигательные сигналы передаются непосредственно от коры к спинному мозгу через кортикоспинальный тракт и опосредованно — через множество вспомогательных путей, вовлекающих базальные ганглии, мозжечок и разные ядра ствола мозга. В целом прямые пути имеют отношение к более изолированным и детальным движениям, особенно кистей и пальцев рук.
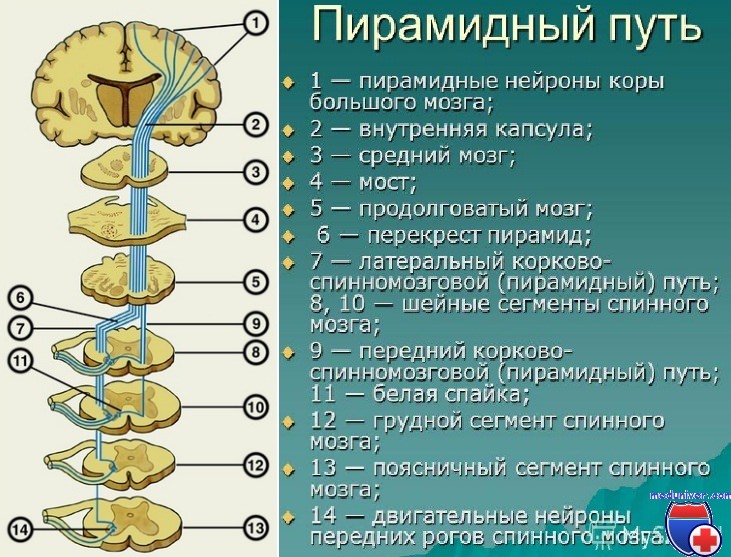
Наиболее важным из путей, выходящих из моторной коры, является показанный на рисунке кортикоспинальный тракт, который называют также пирамидным трактом. Примерно 30% волокон кортикоспинального тракта исходят из первичной моторной коры, 30% — от премоторной и дополнительной моторной коры, а 40% — от соматосенсорных областей, расположенных позади центральной борозды.
После выхода из коры тракт проходит через заднюю ножку внутренней капсулы (между хвостатым ядром и скорлупой базальных ганглиев) и спускается вниз через ствол мозга, формируя пирамиды продолговатого мозга. Основная часть волокон пирамидного тракта в нижней части продолговатого мозга перекрещивается, переходя на противоположную сторону и спускаясь в составе латеральных кортикоспинальных трактов спинного мозга. В основном эти волокна формируют синапсы на вставочных нейронах промежуточных областей серого вещества спинного мозга. Часть волокон заканчиваются на сенсорных релейных (переключающих) нейронах в задних рогах, и очень небольшое число волокон синаптически связаны непосредственно с передними мотонейронами, которые вызывают сокращение мышц.
Небольшая часть кортикоспинальных волокон не переходит на противоположную сторону в продолговатом мозге, а проходит по той же стороне (ипсилатерально) спинного мозга в вентральных кортикоспинальных трактах. Многие из волокон, если не большинство, в итоге тоже переходят на другую сторону спинного мозга на уровне шейных или верхних грудных сегментов. Эти волокна, возможно, связаны с регуляцией билатеральных позных движений, управляемых дополнительной моторной корой.
Наиболее впечатляющей является популяция крупных мякотных волокон пирамидного тракта со средним диаметром 16 мкм. Эти волокна исходят от гигантских пирамидных клеток, называемых клетками Беца, которые обнаруживаются только в первичной моторной коре. Диаметр клеток Беца — примерно 60 мкм, а их волокна проводят импульсы к спинному мозгу со скоростью около 70 м/сек (наибольшая скорость проведения сигналов от головного к спинному мозгу).
В каждом кортикоспинальном тракте примерно 34000 волокон от клеток Беца. Общее число волокон в каждом тракте — более 1 млн, и эти крупные волокна составляют лишь 3% общего числа, а остальные 97% представлены в основном волокнами диметром менее 4 мкм, которые проводят фоновые тонические сигналы в двигательные области спинного мозга.
Другие пути волокон от моторной коры. Моторная кора дает начало большому числу дополнительных, главным образом тонких, волокон, которые идут к глубоким регионам большого мозга и мозгового ствола, включая следующие.
2. Большое число волокон проходит от моторной коры в хвостатое ядро и скорлупу. Отсюда дополнительные пути идут в ствол мозга и спинной мозг, что обсуждается в следующей главе, главным образом для регуляции сокращений постуральных мышц тела.
3. Умеренное количество двигательных волокон идут к красному ядру среднего мозга. От них волокна проходят вниз к спинному мозгу по руброспиналъному тракту.
4. Умеренное число двигательных волокон заходят в ретикулярную формацию и вестибулярные ядра ствола мозга; отсюда сигналы идут к спинному мозгу по ретикуломозжечковым и вестибуломозжечковым трактам.
5. Огромное количество моторных волокон синаптически прерываются в ядрах моста, которые дают начало мостомозжечковым волокнам, несущим сигналы в полушария мозжечка.
6. Коллатерали также заканчиваются в ядрах нижней оливы и отсюда вторичные оливомозжечковые волокна передают сигналы к множественным областям мозжечка.
Таким образом, каждый раз, когда сигнал передается вниз к спинному мозгу для вызова двигательной активности, базальные ганглии, ствол мозга и мозжечок получают мощные сигналы от кортикоспинальной системы.
(trr. corticospinalis anterior et lateral is, пирамидный тракт, корково-спинномозговой тракт, кортико-мускулярный тракт, корково-мышечный тракт)
Латеральный и передний корково-спинномозговые пути проводят нервные импульсы от пирамидных клеток V слоя коры верхних двух третей предцентральных извилин полушарий большого мозга (первичная моторная область - М I, поле 4) и от задних отделов верхней и средней лобных извилин (вторичная моторная область - М II, поля 6, 8) к α-большим мотонейронам передних рогов спинного мозга.
Первые нейроны кортикоспинального тракта представлены гигантскими пирамидными клетками Беца и большими пирамидными клетками V слоя коры. Центральные мотонейроны расположены в коре с учетом соматотопического представительства частей тела на корковый двигательный анализатор. В верхней части моторной области коры, на медиальной поверхности мозга, расположены мотонейроны, обеспечивающие иннервацию поперечно-полосатой мускулатуры контрлатеральной нижней конечности и органов малого таза. Далее, на латеральной поверхности мозга – туловища, верхней конечности (в перевернутом состоянии).
Аксоны первых нейронов входят в состав лучистого венца и идут в нисходящем направлении, в виде компактного пучка направляются во внутреннюю капсулу, занимая передние 2/3 заднего бедра внутренней капсулы.
Далее волокна корково-спинномозгового тракта проходят через середину основания ножки мозга, вентральный отдел моста и на уровне продолговатого мозга по вентральной его поверхности (в основании ствола мозга) формируют пирамиды.
В нижней части продолговатого мозга на границе его со спинным мозгом пирамидный путь разделяется на два пучка: большая часть волокон (75 – 90%) переходит на противоположную сторону, формируя decussatio pyramidum , и располагается после перекреста пирамид в боковых канатиках спинного мозга, образуя латеральный корково-спинномозговой путь (пучок Барнса, Barnes); меньшая часть (около 20%) волокон остается на своей стороне и следует в передних канатиках спинного мозга - передний корково-спинномозговой путь .
Волокна переднего корково-спинномозгового пути, следуя в передних канатиках спинного мозга, посегментно переходят через белую спайку на противоположную сторону и заканчиваются преимущественно в IV – VI пластинах серого вещества на интернейронах.
Особенностью хода переднего корково-спинномозгового пути является частичный перекрест его волокон на уровне сегментов CIII –CIV , где заложены α-большие мотонейроны, обеспечивающие иннервацию мышц диафрагмы, и SIII –SV , где располагаются периферические нейроны для поперечно-полосатой мускулатуры органов малого таза (наружные сфинктеры) .
Аксоны интернейронов передних рогов спинного мозга далее заканчиваются на α-больших мотонейронах, расположенных в IX пластине. Волокна латерального корково-спинномозгового пути в подавляющем большинстве случаев в прямую связь с мотонейронами не вступают (между ними располагаются вставочные нейроны). Импульсы, посылаемые корой, через вставочный нейрон и мотонейрон достигают мышц шеи, туловища, дистальных отделов конечностей.
Следует отметить, что часть аксонов (около 8%) пирамидных клеток вступает в непосредственный контакт с α-большими мотонейронами передних рогов спинного мозга. Особенно много таких прямых контактов с мотонейронами мышц предплечья и кисти, и они характерны преимущественно для переднего корково-спинномозгового пути.
Аксоны вторых нейронов (α-больших мотонейронов) покидают спинной мозг в составе передних корешков, а затем в составе спинномозговых нервов, сплетений стволов периферических нервов и их конечных ветвей направляются к скелетной мускулатуре туловища, шеи и конечностей, заканчиваясь экстрафузально концевой пластинкой нервно-мышечного синапса.
Основная масса пирамидных волокон (около 90%) принадлежит к группе Аd (толщина менее 4 мкм). Группы Аa и Аb (волокна толщиной 11 – 22 мкм) составляют всего около 2% волокон и начинаются от гигантских пирамидных клеток Беца (слой V моторной коры). Таким образом, волокна пирамидного тракта в основной своей массе не являются быстропроводящими – скорость проведения в них составляет всего 25 м/с.
Для корково-спинномозгового тракта соблюдается принцип топической проекции, т. е. каждой точке коры прецентральной извилины соответствует определенная точка (мышца) скелетной мускулатуры. Проекция этого тракта наиболее характерна для дистальной мускулатуры, т. е. для мышц кисти, предплечья (55% волокон), в меньшей степени для мышц стопы и голени (25%), а для проксимальных мышц корковое моторное представительство совсем невелико - 20%. При экспериментальной стимуляции корково-спинального тракта наблюдается отчетливое преобладание активации сгибательной мускулатуры над разгибательной. Более того, в разгибательных мотонейронах обнаруживается тормозной постсинаптический потенциал при стимуляции пирамидного тракта.
Считается, что корково-спинномозговой тракт служит проводником произвольных моторных команд. Однако явное преобладание активации флексорной мускулатуры и торможение экстензоров, наличие посредников в виде интернейронного аппарата при передаче импульса на мотонейрон, а также почти полное отсутствие быстропроводящих толстых миелинизированных волокон заставляют предположить, что корково-спинномозговой тракт не является единственным проводником произвольного движения. Это подтверждается также и тем, что только 40% волокон корково-спинномозгового тракта берут начало в классической моторной области – в 4-м поле Бродмана. Остальные волокна берут начало во вторичной моторной области - задние отделы верхней и средней лобной извилин (поля 6 и 8 Бродмана). Более того, изолированное пересечение кортикоспинального тракта, возможное только в эксперименте на животных, показало отсутствие спастичности и повышения сухожильных рефлексов, что обычно наблюдается в клинических условиях, и даже снижение мышечного тонуса. Это позволило создать представление о тонизирующем действии кортикоспинального тракта на спинальные мотонейроны. В то же время значение корково-спинномозгового тракта в выполнении тонких движений кисти не подвергается сомнению.
КОРТИКОБУЛЬБАРНЫЙ ТРАКТ
(корково-ядерный тракт, tr.corticonuclearis, tr. corticobulbaris)
Корково-ядерный путь - двигательный, нисходящий – представлен цепочкой из двух нейронов и является составной частью пирамидной системы. Этот путь служит для осознанного управления поперечно-полосатой мускулатурой головы и частично шеи, иннервируемой черепными нервами (III, IV, V, VI, VII, IX, X, XI, XII пары). Выполняемые с помощью этого пути движения отличаются исключительной точностью и строгой дифференцировкой.
Корково-ядерный путь начинается крупными пирамидными нейронами, тела которых расположены в V слое коры: в нижней трети прецентральной извилины и заднем отделе средней лобной извилины (корковый цент горизонтального поворота головы и глаз в противоположную сторону). Их аксоны в нисходящем направлении проходят в составе лучистого венца, затем через колено внутренней капсулы, основание средней части ножки мозга, основание моста и достигают центральной части продолговатого мозга.
Надъядерно, на уровне ствола мозга, часть волокон корково-ядерного пути переходит на противоположную сторону, заканчиваясь у клеток двигательных ядер черепных нервов: глазодвигательного (nucl. n. oculomotorius, III), блокового (nucl. n. trochlearis, IV) - в среднем мозге; тройничного (nucl. motorius n. trigemeni, V), отводящего (nucl. n. abducentis, VI), лицевого (nucl. n. faciales, VII)- в мосте; языкоглоточного, блуждающего (nucl. ambiguus- общее для IX, X); добавочного (nucl. ambiguus et nucl. spinalis n. accessorii, XI), подъязычного (nucl. n. hypoglossi, XII)- в продолговатом мозге. Другая часть волокон достигает одноименных ядер своей стороны. Исключение составляют волокна, идущие к участку ядра глазодвигательного нерва (nucl. n. oculomotorius, III), из которого иннервируется внутренняя прямая мышца глаза, к задней порции ядра лицевого нерва (nucl. n. faciales, VII), обеспечивающую иннервацию нижней мимической мускулатуры лица, и к ядру подъязычного нерва (nucl. n.hypoglossi, XII), иннервирующего мышцу языка. Аксоны вторых (периферических) нейронов, тела которых образуют двигательные ядра черепных нервов, направляются в составе соответствующих черепно-мозговых нервов к скелетным мышцам головы и шеи ипсилатеральной (одноименной) стороны.
ТЕКТОСПИНАЛЬНЫЙ ТРАКТ
(tr. tectospinalis, покрышечно-спинномозговой)
Тектоспинальный тракт представляет собой один из древних нисходящих трактов, он начинается от крыши среднего мозга (tectum mesencephali) - нейронов верхних бугров четверохолмия (colliculus superior). Возможно, что нижние бугры также участвуют в формировании этого тракта, но морфологически это пока не подтверждено. Вентрально от водопровода мозга (aqueductus cerebri) тектоспинальные волокна перекрещиваются (фонтановидный перекрест Мейнерта, Meynert) и спускаются в передние столбы спинного мозга, располагаясь вдоль передней срединной его щели, медиальнее переднего пирамидного тракта. Существует также и боковой тектоспинальный тракт, который проходит в боковых канатиках спинного мозга. Проекции тектоспинального тракта осуществляются на мотонейроны проксимальных мышц-разгибателей и мышц туловища до уровня C IV , причем эта проекция перекрывается в спинном мозге с проекцией руброспинального тракта.
Физиологическая функция тектоспинального тракта, вероятнее всего, заключается в обеспечении координированных движений глаз, головы и верхних конечностей на неожиданные световые и звуковые воздействия. Наличие спинно-тектального сенсорного тракта подтверждает подобную точку зрения. При экспериментальном пересечении тектоспинального тракта происходит резкое замедление аудио- и видео спинальных реакций.
Аналогом тектоспинального тракта для двигательных ядер ствола мозга является тектобульбарный тракт (пучок Мюнцера, fasc. Munzeri) .
ТЕГМЕНТО-СПИНАЛЬНЫЙ ТРАКТ
Тегменто-спинальный тракт начинается от покрышки среднего мозга и нисходит до уровня С IV спинного мозга. Возбуждение волокон тракта приводит к уменьшению поступления афферентной информации от мышечных веретен мышц-разгибателей. Таким образом, торможение разгибателей приводит к преобладанию тонуса сгибательной мускулатуры. Поэтому функцию тегменто-спинального тракта связывают с выработкой позных оборонительных и ориентировочных реакций.
ОЛИВО-СПИНАЛЬНЫЙ ТРАКТ
(tr. olivospinalis, пучок Гельвега, fasc. Helwegi,
биологические науки
- Ланская Елена Владимировна , аспирант
- Великолукская государственная академия физической культуры и спорта
- МЕТОДЫ ОЦЕНКИ ФУНКЦИОНАЛЬНЫХ СВОЙСТВ МОТОРНОЙ СИСТЕМЫ
- КОРТИКО-СПИНАЛЬНЫЙ ТРАКТ
- РЕГУЛЯЦИЯ ДВИЖЕНИЙ
- УПРАВЛЕНИЕ ДВИЖЕНИЕМ
Похожие материалы
- Изучение электронейромиографических параметров вызванных потенциалов мышц верхних и нижних конечностей у представителей видов спорта с близкой по структуре двигательной деятельностью
- Параметры моторных ответов мышц бедра, голени и стопы у представителей различных видов спорта при магнитной и электрической стимуляции спинного мозга и периферических нервов
- Амплитудно-частотные характеристики электромиографической активности скелетных мышц у пауэрлифтеров при выполнении упражнений силового троеборья
- Применение электро- и психофизиологических методов в спортивной физиологии и для оценки эффективности физической реабилитации лиц с отклонениями в состоянии здоровья
- Исследования почвенной микрофлоры
Вопрос контроля движений является кардинальным для понимания природы и механизмов двигательного поведения человека. Известна нисходящая управляющая роль двигательных отделов головного мозга в согласовании работы мышц через соответствующие моторные нейрональные контуры спинного мозга [1]. Выделяются две группы нисходящих путей от супраспинальных структур: медиальная и латеральная. Медиальная группа включает вестибуло-спинальный и ретикуло-спинальный тракты, которые иннервируют мышцы туловища, плечевого и тазового поясов, обеспечивают регуляцию мышечного тонуса и осуществление позотонических реакций. К латеральной группе относят кортико-спинальный и кортико-красноядерно-спинальный тракты, которые в основном иннервируют мышцы дистальных отделов конечностей и обеспечивают выполнение тонких движений кисти и стопы [2].
Двигательные корковые поля имеют прямые связи с МН и вставочными нейронами спинного мозга через кортико-спинальный тракт и поддерживают непрямые контакты с нижними нейронами через нисходящие пути ствола головного мозга [5], особенно с помощью кортико-ретикулоспинальных проекций. Нейроны 4-го поля контролируют тонкие произвольные движения скелетных мышц противоположной половины тела, поскольку большинство волокон пирамидного тракта переходит на противоположную сторону в нижней части продолговатого мозга. Стимуляция поля 4 индуцирует общие движения отдельных мышц, в то время как стимуляция поля 6 вызывает более сложные движения, такие как движения всей руки или ноги [9]. В ряде работ указывается, поле 6 рассматривалось как дополнительное моторное представительство мускулатуры туловища, но в последнее время получены данные о его ведущей роли в качестве ассоциативного поля [10], участвующего в подготовке и реализации движения, особенно если ранее оно было разучено [11].
Импульсы пирамидных клеток двигательной коры идут по двум путям, заключенным в ростральной части пирамидного тракта. Один из них – корково-ядерный путь, который заканчивается на ядрах черепных двигательных нервов в стволе мозга. Другой – кортико-спинальный путь, который оканчивается на вставочных нейронах спинного мозга, которые, в свою очередь, синаптически связаны с большими МН передних рогов. Эти нейроны передают импульсы по передним корешкам спинного мозга и периферическим нервам к двигательным концевым пластинкам скелетных мышц [9].
Покинув двигательную кору, волокна кортико-спинального пути проходят через белое вещество в составе лучистого венца и сходятся в области заднего бедра внутренней капсулы. В очень компактном виде они проходят в соматотопическом порядке внутреннюю капсулу и вступают в среднюю часть ножек мозга. Здесь они представляют собой компактный пучок, который опускается в центр основания каждой из половин моста, будучи окруженным множеством нейронов ядер моста и волокнами различных систем. На уровне перехода моста в продолговатый мозг пирамидные пути становятся заметными снаружи и формируют удлиненные перевернутые пирамиды по обе стороны от передней средней линии [12]. На границе продолговатого и спинного мозга большая часть кортико-спинальных волокон (до 80%) переходит на противоположную сторону (волокна ипсилатерального кортико-спинального тракта), а меньшая продолжает идти по своей стороне. В результате в спинном мозге формируются два кортико-спинальных пути. Основным является перекрещенный латеральный кортико-спинальный путь, идущий в боковом канатике. Неперекрещенный прямой кортико-спинальный путь проходит в переднем канатике спинного мозга. На уровне каждого сегмента волокна как прямого, так и латерального кортико-спинальных путей заканчиваются у МН передних рогов. Показано, что оба кортико-спинальных тракта являются глутаматергическими, оказывая возбуждающее влияние моно- и полисинаптически на альфа-мотонейроны (α-МН) и полисинаптически – на гамма-мотонейроны (γ-МН) [6]. Прямой кортико-спинальный путь обеспечивает проведение нервного импульса к МН грудных сегментов, иннервирующих мышцы туловища. Через латеральный кортико-спинальный путь импульсы поступают к МН всех (в том числе и грудных) сегментов. Таким образом, мышцы конечностей получают одностороннюю корковую иннервацию от прецентральной извилины противоположного полушария, а мышцы туловища обеспечены двусторонней корковой иннервацией. Этим объясняется тот факт, что при одностороннем поражении кортико-спинального пути в головном мозге (выше перекреста) расстройства произвольных движений возникают в мышцах конечностей на противоположной стороне. Одностороннее поражение кортико-спинального пути в спинном мозге (ниже перекреста) приводит к расстройствам произвольных движений на стороне поражения. Двигательные расстройства в мышцах возможны, но лишь при условии их двустороннего поражения [7].
Волокна ипсилатерального тракта берут начало преимущественно от дополнительных моторных областей, включая поле 6. Характер расположения нейронов ипсилатерального пути объясняет, почему методически при стимуляции поля 4 ипси-ВМО не регистрируется [12]. До сих пор ведутся споры по поводу того, насколько волокна ипсилатерального кортико-спинального тракта участвуют в генерации ипсилатерального ВМО, регистрируемого в норме и при патологии в ответ на транскраниальную электрическую стимуляцию (ТЭС) и транскраниальную магнитную стимуляцию (ТМС) [13]. Не исключено, что в реализации ипсилатерального ВМО основную роль играют кортико-ретикулоспинальные пути [14].
Интернейроны, входящие в состав полисинаптической цепи, играют первостепенную роль в определении характера импульса к МН – конечному общему пути. В связи с этим интернейроны могут рассматриваться по отношению к МН как еще один, более высокий уровень регуляции его активности. Интернейроны при этом получают импульсацию не только от двигательной коры, ствола мозга и лимбической системы, но и от афферентных периферических нервов. Эти клетки согласуют интегральные действия мышц-синергистов и тормозят работу мышц-антагонистов, регулируя, таким образом, их совместную деятельность, контролируют защитные и позные рефлексы, опосредуют стереотипные поведенческие реакции, а также в определенной степени обеспечивают реализацию высокодифференцированных движений. Интернейроны могут также контролировать контрлатеральный моторный пул. Разновидность интернейронов – клетки Реншоу, которые регулируют возбудимость α-МН с помощью отрицательной обратной связи, используя такие тормозные нейротрансмиттеры, как глицин и, возможно, таурин. Таким образом, мышечные сокращения – это конечный результат сложной иерархической организованной системы двигательного контроля. Изучение этого контроля важно для понимания механизмов формирования двигательного акта в норме и при патологии [6].
Окончательное суждение о функции кортико-спинального тракта у человека по результатам морфологических исследований и клинико-морфологических сопоставлений пока невозможно. Вместе с тем существует общее мнение, что кортико-спинальный тракт обеспечивает не только эфферентную функцию - организацию движения и регуляцию мышечного тонуса, но и афферентную – поступление информации в кору, подкорковые структуры и мозжечок от периферических сенсорных рецепторов и спинальных МН [6; 8]. Были сформированы гипотезы о роли кортико-спинального тракта в деятельности ЦНС. В частности, в обзоре R.A. Davidoff [15] обсуждаются три основных положения. Первое положение: кортико-спинальный тракт занимает центральную позицию в конвергенции деятельности различных популяций корковых нейронов, суммируя информацию посредством прямых моносинаптических связей с большими клетками (α-МН) передних рогов спинного мозга. При этом переключающиеся в подкорковых структурах кортико-фугальные (по-другому нисходящие, эфферентные) волокна действуют параллельно с моносинаптическими. Следует отметить, что моносинаптическая природа кортико-спинальных проекций показана для дистальных мышц кисти, мышц лица и языка, мышц проксимальных отделов рук (дельтовидная мышца) и плечевого пояса (трапециевидная и большая грудная мышцы), а также мышц ног [16]. Второе положение: кортико-спинальный тракт особенно важен в контроле тонких и раздельных движений пальцев руки, причем в реализации этого акта участвуют как моносинаптические, так и полисинаптические волокна кортико-спинального тракта [15]. Третье положение: кортико-спинальный тракт представляет собой путь для кортико-фугальной регуляции различных сенсорных переключений, Этот контроль сенсорного входа в центральные структуры может влиять на тип и объем соматосенсорной и проприоцептивной информации, поступающей в сенсорную и ассоциативную кору [15].
Использование современных нейрофизиологических методов и, в частности, ТМС позволяет не только оценивать верхнюю и нижнюю составляющие кортико-спинального тракта, но также анализировать функциональные взаимоотношения корковых нейронов и сегментарных рефлекторных контуров, активируемых при участии кортико-спинальных путей [3; 17; 18; 19]. Корковая стимуляция с использованием переменного магнитного поля является одной из самых физиологичных методик, так как показано, что ТМС возбуждает именно те корковые нейроны, которые первыми активируются при совершении произвольного движения [12; 20; 21]. Показано, что ТМС может быть использована для оценки изменений функциональных свойств моторной системы человека под влиянием мышечной работы различной направленности 26. В отличие от ТМС, позволяющей изучать исключительно механизмы функционирования верхней составляющей кортико-спинального тракта, магнитная и электрическая стимуляция спинного мозга, в свою очередь, дают возможность исследовать его нижнюю составляющую и наряду с методами электро- и магнитной стимуляции периферических нервов оценивать функциональное состояние его периферической части как в норме (с участием представителей различных видов спорта и нетренированных лиц), так и при патологии 37. Использование такого арсенала методов в практике спорта обеспечивает изучение особенностей нейропластичности головного и спинного мозга под влиянием специфической мышечной деятельности и функционирования соответствующих периферических звеньев нервно-мышечного аппарата и способствует выявлению механизмов развития адаптационных процессов в нейромоторной системе лиц, длительное время регулярно выполняющих физическую нагрузку.
Список литературы
Электронное периодическое издание зарегистрировано в Федеральной службе по надзору в сфере связи, информационных технологий и массовых коммуникаций (Роскомнадзор), свидетельство о регистрации СМИ — ЭЛ № ФС77-41429 от 23.07.2010 г.
Соучредители СМИ: Долганов А.А., Майоров Е.В.
Будет сборник постов по теме организации спинного мозга

Содержание
- 1 Тракты спинного мозга
- 1.1 Куйперс и целевые зоны спинного мозга
- 2 Кортикоспинальный тракт
- 2.1 Лемон и функции кортикоспинального тракта
- 2.2 Олег о прорастании и связи с гипотонией
- 2.3 Сарнат и функции кортикоспинального и кортикобульбарного трактов в развитии
- 2.4 Толщина аксонов и скорость проводимости в кортикоспинальном тракте
Тракты спинного мозга

Для иллюстрации мысли о разных вкладах в спастические состояния, и для раскрытия темы ретикулоспинальных против вестибулоспинальных построений приведу картинку этих трактов на уровне где-то верхних шейных позвонков Красным — нисходящие пути Обращаем внимание, что кортикоспинальный и ретикулоспинальный тракты имеют две компоненты в спинном мозгу (fasciculi) - латеральный и медиальный
Заметим (повторяясь) что кроме экстрапирамидальных и пирамидальных классификаций взгляд на эту картину подсказывает и группировку пучков латеральные(боковые) - медиальные(Срединные). Они не только проходят в общих пучках, но и прорастают в близкие места спинного мозга потом. Ретикулоспинальный и кортикоспинальный тракты присутствуют и там и там.

Очень поучительная схема, адаптированная из Куйперса (60е годы). Показано, в какие зоны приходят нисходящие, эфферентные пути. Слева — пирамидальные, справа — экстрапирамидальные. И очень четко видно, что только первичная моторная кора, и лишь в малой своей части приходит в передний рог серого вещества спинного мозга, непосредственно к моторным нейронам. Все остальные пути выходят к интернейронам: и контролируют не мышцы, а "готовые" спинальные механизмы: рефлексы, автоматизмы и т. п. Считается, что именно эти прямые пути управляются огромными клетками им. Владимира Беца. У приматов-нечеловеков эта доля прямых соединений еще меньше, у кошек и пр. — еще меньше.
Та же мысль, тоже из Куйперса, но цитируемая по Альтману, книжка про спинной мозг: (А) - кошка, (B) - макака, (С) - шимпанзе. Показаны кортикоспинальные терминалы в переднем роге спинного мозга на уровне шейного отдела. У кота - почти ничего, а у шимпанзе - сильная концентраия в латеральных моторных колоннах, там где пальцы и кисть

Кортикоспинальный тракт

Приведу функции кортикоспинального тракта по прекрасному обзору Лемона (который рекомендую) 2008 года по моторным цепям — в качестве независимого от Sarnat изложения темы. У него практически нет аспекта неонатального развития, зато есть компаративный
Итак, функции кортикоспинального тракта:
- Супраспинальный контроль сенсорных (включая болевые) цепей
- выбор, управление и контроль силы (gain control) спинальных рефлексов
- Прямое и косвенное возбуждение и торможение мотонейронов
- Управление автономной нервной системой
- "обучение" спинальных нервных систем
- во время развития — трофические функции, то есть управление прорастанием нервных окончаний
В связи с тем, что тема гипотонических симптомов получила развитие и идея гиперэластичности имеет своих сторонников, опишу еще раз, что имею в виду:
- Сегмент спинного мозга формируется в эмбриональный период.
- В момент формирования к нему прорастает кортикоспинальный тракт.
- Дозревание сегмента происходит под влиянием кортикоспинального тракта.
- Кортикоспинальный тракт подходит не только к передним рогам спинного мозга, но и к интернейронам.
- Нарушение сопряжения кортикоспинального тракта с спинальным сегментами приводит к картине локальной гипотонии.
- Гипотонический синдром может сопровождать и картину спастического тетрапареза и синдрома, который ранее назывался минимальной мозговой дисфункцией.
Нет смысла искать нарушение созревания эластических, коллагеновых или фибриновых волокон в столь большом количестве случаев. Альтернативой этому суждению может быть признание необычайной распространенности генетических болезней нарушения формирования эластических, коллагеновых и другого рода волокон. Мысль о том, что спиномозговой сегмент может быть "открыт" для формирующих влияний, плохо проходит из-за слабой визуализации (визуального образа) этого процесса. Мне кажется, что большая часть читателей представляет, что сегмент сразу принимает окончательный вид и нет периода, когда прорастающий к нему кортикоспинальный тракт входит в него, фиксируется в определенном месте и начинает проводить кортикальные влияния.
Программное ревью Sarnat, которое отражает точку зрения ведущего детского невролога на становление произвольности и того, почему при повреждении в перинатальном периоде проявления выглядят так, а не иначе - Олег К


Ktyz Vbiby в недрах дискуссии в соседнем посте сказал "Со скоростями проводимости вообще темный лес. Там есть аномалии, при которых скорость втрое ниже нормы, но выявляются эти люди случайно. То есть полная норма. Приятельница миографистка рассказывала, что ей такой чемпион Москвы попался по велогонкам"
- Заметим так же что миографисты меряют падение скорости проводимости, происходящее при некоторых заболеваниях, и говорят, что люди надежно мрут уже при скоростях в пару раз выше чем та, с которой благополучно живут упомянутые аномальные. (добавил Ktyz Vbiby).
Это действительно факт, который при чтении текстов учебников меня тоже сильно беспокоит. Потому как игнорируют они его.
Вот график распределения толщины аксонов в кортикоспинальном тракте. Это как бы тот самый тракт их "первичной" моторной коры к мотонейронам спинного мозга, про который учебники дают понять, что он и рулит движениями, чуть ли не напрямую. Особенно раздельными движениями пальцев.
Однако 70% аксонов в нем имеют толщину в 1μm, и скорости проводимости, соответственно 3-4 метра в секунду. Т.е. сигнал до даже до шейных отделов позвоночника будет добираться ощутимые 0.1 сек, что кажется невероятно плохо для "управления" быстрыми действиями.
И только жалкие 0.4% аксонов имеют миелиновую оболочку больше 10 μm и скорости 80 метров в секунду и более.
Кто бы не "проектировал" этот тракт, скорость явно не была у него в приоритете. Скорость явно не критична для работы абсолютного большинства его составляющих.
При этом глядя на игру "раздельными движениями пальцев", например, Глена Гульда, который мало того что быстр, но и мастерски окрашивает каждый звук им извлекаемый — остается только поражаться, как же он этого достигает.
P.S. График слепенький, потому что более свежего, чем вот этот из статьи 1970г я не нашел. На него до сих пор ссылаются (и на более ранние работы), а новых — не делают. Было б, скажем, интересно проследить, куда ходят эти 0.4% Это ж 5000 волокон "всего". У макак, на которых их трассируют, и того меньше.
Откуда они идут и куда?
Вот кстати (справа) еще интересная иллюстрация из все той же работы группы Лемона по структуре кортикоспинального тракта.
Показано распределение задержек в зависимости от источника аксонов — моторная кора - M1 и две премоторные — SMA и F5 (медиальная и латеральная премоторные области).
М1 очень быстрые, премоторные — гораздо медленней, особенно F5
Читайте также:
